









Introduction
Oxidative damage generally occurs due to free radicals and reactive oxygen species. Synthetic antioxidants (e.g., propyl gallate, butylated hydroxytoluene, and butylated hydroxylanisole) are commonly used to act against free radicals in biological systems and food (Bouayed and Bohn, 2010). However, the use of these radical scavengers is restricted due to potential health risks (Chi et al., 2014; Zarei et al., 2014). Inflammation is a major defense mechanism against pathogens and is stimulated by a range of microbial products such as lipopolysaccharides (LPS). Macrophages exhibit a particularly vigorous response to LPS, and macrophage activation by LPS enhances the production and release of inflammatory mediators such as tumor necrosis factor-α (TNF-α), a pro-inflammatory cytokine (Guastadisegni et al., 2002). However, the production of excessive inflammatory cytokines must be regulated, since it can lead to harmful inflammatory responses such as rheumatoid arthritis, septic shock, and other chronic inflammatory diseases (Kim et al., 2011).
Pork is one of the most popular meats in Korea, and the level of pork consumption has increased in recent years. Pork consumption per capita in 2005 was 17.8 kg and rose to 20.89 kg in 2013 (Korea Meat Trade Association, 2015). Pork cuts containing high fat content such as the belly and Boston butt are popular, and low fat portions such as loin and ham are less preferred by Korean. However, consumers have recently had a negative preconception about livestock products (Cross et al., 2007). They misunderstand that meats contain high amo-unts of fat and believe this is a causative factor for the risk of obesity, hypertension, cardiovascular disease, cancer, etc. (Özvural and Vural, 2008). However, meat is a good source of protein with high biological value. Especially, pork is an excellent source of valuable nutrients such as vitamins, minerals (e.g., vitamin B12, folic acid, and iron), and antioxidative peptides. Histidine-di-peptides such as carnosine (β-alanyl-L-histidine) and anserine (N-β-alanyl-3-methyl-L-histidine) are functional peptides with antioxidative activities and are widely distributed in skeletal muscle, the central nervous system, and the heart at very high concentrations (up to 20 mM) (Quinn et al., 1992). These antioxidant di-peptides are obtained from the muscle of animals such as pork, beef, chicken, and fish. Their wellknown functions include discarding toxins from the body and regulating the immune function of macrophages (Guiotto et al., 2005). In our previous study, the carnosine content of raw cuts of loin and ham from Korean native black pigs (KNBP) was determined to be 1.35 mg/mL and 1.27 mg/mL, respectively (Kim et al., 2013). Although pork has many nutritional benefits, very few scientific studies have examined its antioxidation and anti-inflammation effect based on different cuts. Also, pork is often consumed by various cooking methods such as grilling, boiling, and stewing. Many water soluble compounds in pork can be eluted in the hot water when it cooked. Therefore, the objective of this study was to investigate the effects of boiled pork and hot water extracts of four different cuts on radical scavenging activity and proinflammation cytokine reduction activity.
Materials and Methods
Four different pork cuts such as loin, boston butt, shoulder ham, and ham of LYD (castrated, n=4) were purchased from a local market in Chuncheon. All subcutaneous, intermuscular, and visible connective fat were removed and then each raw meat cut was added to five-fold distilled water and boiled for 1 h and filtered with cheese cloth (Fig. 1). The boiled pork and hot water extracts were separated and lyophilized. The lyophilized powder was dissolved in distilled water according to the concentration and used as sample for further analysis.
Carnosine and anserine content were analyzed in accordance with the methods described by Gil-Agusti et al. (2008). A 3 g sample combined with 15 mL distilled water was homogenized (Polytron PT-2500 E, Kinematica, Switzerland) for 10 min at 15,000 rpm. Homogenized samples were centrifuged at 3,000 rpm for 10 min. The slurry was filtered through Whatman No. 1 filter paper. The filtrate was combined with chloroform, shaken for 30 s, and then centrifuged at 3,000 rpm at 4℃ for 10 min. The supernatant was used for dipeptide content analysis. The samples were conducted using an HPLC (Agilent Technologies 1260 Infinity, USA) and a zorbax eclipse XDB-C18 column (250×4.6 mm, 5 μm, Agilent, USA). The samples were analyzed at a flow rate of 1 mL/min for 20 min, column temperature of 25℃, and UV absorbance at 210 nm. The mobile phase was comprised of 0.01 M sodium dodecyl sulfate soluble in 0.01 M sodium dihydrogen phosphate at pH 7.0, and sodium hydroxide was used for validation. Sample volumes injected were 20 μL. All samples and solvents were filtered through 0.45 μm membrane filters. Standard carnosine and anserine were purchased from Sigma (USA). The experimental values are indicated linearity, using the area ratio method.
The DPPH radical scavenging activity of BPP and HWEP were measured in accordance with Blois (1958), with slight modification. Briefly, 0.2 mM DPPH solution was mixed with each concentration sample in distilled water after which the mixture was vortexed and incubated at room temperature for 30 min. After standing for 30 min, the absorbance was determined at 517 nm using a UV/VIS spectrophotometer (SpectraMax M2e, Molecular Devices, USA) and calculated as millimolar Trolox equivalents (TE) per gram of low molecular weight sample.
ABTS+ scavenging activity was carried out according to the procedure described by Re et al. (1999). ABTS+ was produced by reacting 14 mM ABTS treated with 4.9 mM potassium persulfate. Prior to use, the mixture was vortexed and incubated in the dark at room temperature for 12-16 h to generate a dark blue solution. The ABTS+ solution (SpectraMax M2e, Molecular Devices, USA) was diluted in 5.5 mM PBS (pH 7.4) and equilibrated to 30℃ until the absorbance reached 0.70±0.02 at 734 nm. The reactions consisted of 50 μL sample and 950 μL of the ABTS+ radical solution. Absorbance was measured (SpectraMax M2e, Molecular Devices, USA) at 30℃ after 30 min and calculated as millimolar TE per gram of low molecular weight sample.
The capability of reducing ferric ions was assessed according to the Benzie and Strain (1996) method, with some modification. The FRAP reagent was produced and added to acetate buffer (300 mM/L, pH 3.6), 2,4,6-tripyridyl-S-triazine (TPTZ, 10 mM in 40 mM/HCl) solution, and FeCl3·6H2O (20 mM) at a ratio of 10:1:1 (v/v). The mixture was incubated at 37℃. To test tubes, 25 L of sample and 175 L of FRAP reagent were added and incubated at 37℃ for 5 min. The absorbance of FRAP was recorded at 595 nm (SpectraMax M2e, Molecular Devices, USA). The FRAP assay for each sample was expressed as mol TE per g of weight and calculated from the linear calibration curve.
The oxygen radical absorbance activity assay was performed using a modified version of Gillespie et al. (2007). Containers with 150 L of fluorescein (80 nM) and 25 L of sample were shaken, and each well contained a final volume of 200 μL solution. The mixture was placed at 37℃ for 15 min. Then, 25 L of AAPH (150 mM) was added to each well. A fluorometric spectrophotometer (SpectraMax M2e, Molecular Devices, USA) was used to record absorbances (excitation wavelengths of 480 and 520 nm, intervals for 60 min) after each set was automatically shaken at 37℃. Trolox standards and a blank sample were also used. The ORAC values were calculate using the respective area under curve (AUC) and regression equations between the TE and the net AUC. The final ORAC values are expressed as mol of TE.
A mouse macrophage cell line, RAW 264.7, was obtained from American Type Culture Collection (ATCC TIB-71TM, USA). RAW 264.7 cells were plated in 100mm tissue culture dishes containing Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (Welgene, USA) and 1% penicillin/streptomycin (Gibco, USA). RAW 264.7 cells were maintained at 37℃ in a humidied incubator containing 5% CO2.
RAW 264.7 cells were plated at a density 5×104 cells/mL in tissue culture plates and incubated for 24 h. Five microliters of the sample with various concentrations (10, 50, 100, 200, and 500 μg/mL) was transferred to the wells. After an hour, the cells were stimulated with LPS (1 μg/mL) and incubated for 24 h at 37℃. To each well, 5 mg/mL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) (50 μL) was added and incubated for 4 h at 37℃. The medium with MTT was removed and then dissolved in dimethyl sulfoxide (DMSO). The optical density (OD) of each well was measured at 540 nm (SpectraMax M2e, Molecular Devices, USA).
Cells were plated density 2×105 cells/mL and grown to confluence in 24-well tissue culture dishes. Cells were cultured in an incubator (37℃ and 5% CO2) for 24 h. On the following day, the medium was replaced with fresh serum free medium and the wells were loaded with 5 L of sample at various concentrations (10, 50, 100, 200, and 500 μg/mL). After approximately 1 h, the medium was stimulated with LPS (1 μg/mL) and incubated for 24 h at 37℃. The medium with LPS was collected and centrifuged at 12,000 rpm for 5 min at 4℃. The levels of IL-6 and TNF-α were analyzed by ELISA as per the manufacturer’s instructions. IL-6 and TNF-α were determined using mouse IL-6 BD OptEIATM, ELISA sets (BD Phar- Mingen, USA) and mouse TNF (Mono/Mono) BD OptEIATM, ELISA sets (BD PharMingen, USA), respectively. The coating buffer contained capture antibody placed in a 96-well plate and incubated overnight (approximately 16 h) at 4℃. The capture antibodies were aspirated and washed 3 times. Blocking solution (Assay Diluent) was added and incubated for 1 h at room temperature. Standards (15.6, 31.3, 62.5, 125, 250, 500, and 1,000 pg/mL) or samples were added to wells and incubated for 2 h at room temperature. The standards and samples were removed and washed 6 times. Working Detector (Detection Antibody + SAv-HRP reagent) was added to each well and incubated for 1 h, followed by washing 6 times. To each well, 100 L of substrate solution (tetramethylbenzidine and hydrogen peroxide) was added. The plate was incubated for 30 min at room temperature in the dark. The stop solution (2 N H2SO4) was added to each well and absorbance from 450-540 nm was read.
All results were analyzed using one-way analysis of variance (ANOVA) according to the general linear model procedures for SAS software (ver. 9. SAS Institute Inc., USA). Results are shown as mean values and standard error of the sample. Duncan's multiple range tests were used to determine statistical significance between the treatments (p<0.05).
Results and Discussion
Table 1 shows the carnosine and anserine content of BPP and HWEP from the four pork cuts. In the case of carnosine, both BPP and HWEP of loin resulted in significantly higher carnosine content compared with the other pork cuts. Particularly, carnosine in HWEP of the four cuts was higher than BPP. The anserine content of both BPP and HWEP in shoulder ham was higher than that of BPP and HWEP of the other pork cuts. Tinberge and Slump (1976) reported that the carnosine and anserine concentration of pork was 104-338 and 7-16 mg/100 g, respectively. Aristoy and Toldrá (1998) suggested that the carnosine content of four pork muscles (masseter, trapezius, semimembranosus, and longissimus dorsi) were 21.06, 180.98, 320.84, and 313.02 mg/100 g muscle, respectively. Also, anserine contents of those muscles were 6.09, 10.68, 17.56, and 14.56 mg/100g muscle. They suggested that the caronsine and anserine levels are a good indication of muscle glycolytic activity. Lumber longissimus dorsi (loin) and semimembranosus (ham) were classified as glycolytic muscle due to higher carnosine content. In cooked beef, the carnosine content was 134 mg/100 g of tissue (Park et al., 2005). The higher fat content of beef could have a low concentration of carnosine, since carnosine is mainly present in the cytosol of skeletal muscle and the higher adipose fat would decrease skeletal muscle and thus carnosine concentration (Park et al., 2005). Crush (1970) reported that the carnosine and anserine concentration of beef leg muscle was 150 and 50 mg/100 g of tissue, respectively. In addition, Plowman and Close (1988) reported that the carnosine and anserine concentrations in beef shoulder and rib steak were 341 and 58 mg/100 g of tissue, respectively. These results suggested that animal muscles have different carnosine content by classification if it is glycolytic or oxidative muscle.
A-DMeans±S.E. within same row with different superscript letters differ significantly at p<0.05.
a-dMeans±S.E. within same column with different superscript letters differ significantly at p<0.05.
1)CNS, carnosine; ANS, anserine, BPP, boiled pork powder; HWEP, hot water extracts powder.
The antioxidative effects, mediated by various radical scavenging mechanisms, including DPPH, ABTS radical scavenging activity, FRAP, and ORAC of BPP and HWEP of LYD are shown in Table 2. These assays are well known as screening methods for determining the total antioxidant capacity of foods and beverages (Alvarez-Suarez et al., 2012). BPP of Boston butt exhibited the highest DPPH radical scavenging effect (13.65 M TE) compared with the other pork cuts. However, HWEP of loin (19.40 M TE), shoulder ham (16.96 M TE), and ham (21.45 M TE) exhibited significantly higher DPPH radical scavenging effects compared with Boston butt. The ABTS radical scavenging activity of BPP and HWEP of the four pork cuts are shown in Table 2. BPP of shoulder ham and ham exhibited significantly higher ABTS scavenging activity, 39.28 and 39.43 M TE, respectively. However, HWEP of ham had the highest ABTS radical scavenging effect at 43.96 M TE compared with that of the other pork cuts. In this study, we found that the ABTS radical scavenging effects of BPP and HWEP were a slightly higher than the DPPH radical scavenging activities. This may be because the scavenging activity of the DPPH radical assay is only suitable for lipid-soluble antioxidants and for the evaluation of hydrogen-donation of antioxidative compounds, whereas the scavenging activity of the ABTS radical assay assesses the single electron-transfer capabilities of these compounds and is applicable to both water-soluble and lipid-soluble antioxidants (Re et al., 1999; Serpen et al., 2012). Therefore, the measured ABTS radical scavenging activity values were higher than the measured DPPH radical scavenging results, and this might reflect various aspects of their antioxidant properties and different chemical mechanisms (Serpen et al., 2012).
A-DMeans±S.E. within same row with different superscript letters differ significantly at p<0.05.
a-dMeans±S.E. within same column with different superscript letters differ significantly at p<0.05.
1)Sample concentration for antioxidative activity was 1 mg/mL.
2)BPP, boiled pork powder, HWEP, hot water extract powder.
The results of the FRAP assay for HWEP from ham appeared to have a similar trend compared to the ABTS radical scavenging activity, although no significant difference was found between that of shoulder ham and ham. Serpen et al. (2012) reported that the FRAP value of raw meat samples (beef tenderloin, chicken breast, pork tenderloin, and sea bream fish fillet) were between 3.0 and 4.9 mmol TE/kg. In addition, they indicated that beef samples had the highest FRAP value (4.9±0.2 mmol TE/kg), whereas the fish samples had the lowest (3.0±0.1 mmol TE/kg).
The antioxidative effects of BPP and HWEP from four different pork cuts were obtained using the ORAC assay (Table 4). The HWEP of loin, Boston butt, and ham generally had high ORAC values. Heat treatment might increase the antioxidant activity of some proteins due to structural changes. In this study, we have found that HWEP had higher ORAC values than BPP from loin, Boston butt, shoulder ham, and ham. In this study, we have found that the HWEP showed more high antioxidative activity compare to BPP (p<0.05). We assumed that the high antioxidative activity of the HWEP from pork cuts such as loin and ham might due to antioxidative peptides carnosine. Because it is a very important natural water soluble muscle di-peptide and it was eluted in hot water from the pork during heating. Also it can contribute to the inactivation of lipid oxidation catalysts and/or free radicals in the sarcoplasm (Decker and Crum, 1993). Also, denaturation of the proteins can increase their ability to scavenge radicals by increasing the solvent exposure of antioxidant amino acids (Serpen et al., 2012). Wu et al. (2008) examined ORAC in beef extracts and consequently determined that beef extracts prepared with low ethanol concentrations (20%) demonstrated higher hydrophilic ORAC. In addition, it has been demonstrated that proteins and peptides have an important antioxidant action in meats due to their ability to scavenge free radicals. Soy protein, similar to meat, is potential antioxidant, and antioxidant peptides obtained from soybean protein have been reported in a number of studies (Hernndez -Ledesma et al., 2009).
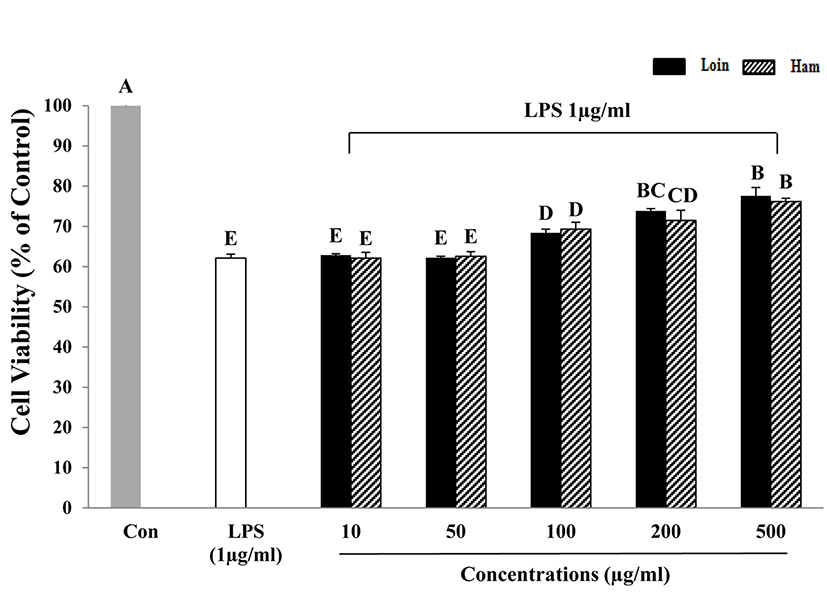
To investigate the anti-inflammatory effects of HWEP from loin and ham on macrophage cells (RAW 264.7), 1 μg/mL of LPS was used for treatment. The LPS-induced RAW 264.7 cells were treated with various concentrations of HWEP (10, 50, 100, 200, 500 μg/mL). Cell viability of LPS-induced RAW 264.7 cell was increased with increased HWEP concentration. HWEP concentrations more than 100 μg/mL resulted in significantly higher cell viability compared with LPS-induced RAW 264.7 cells alone. LPS-induced cells without HWEP treatment exhibited a cell viability of 61-63%, which was approximately 40% lower than control cells.
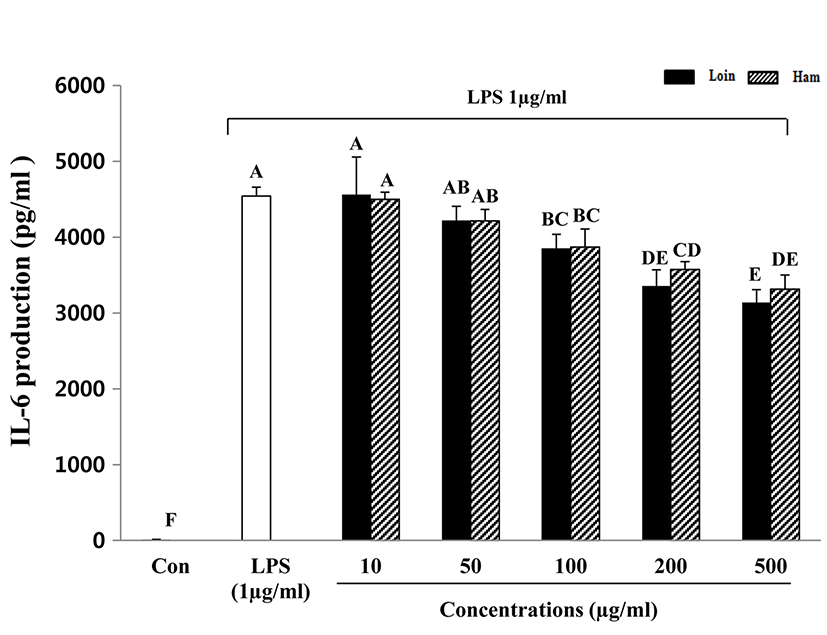
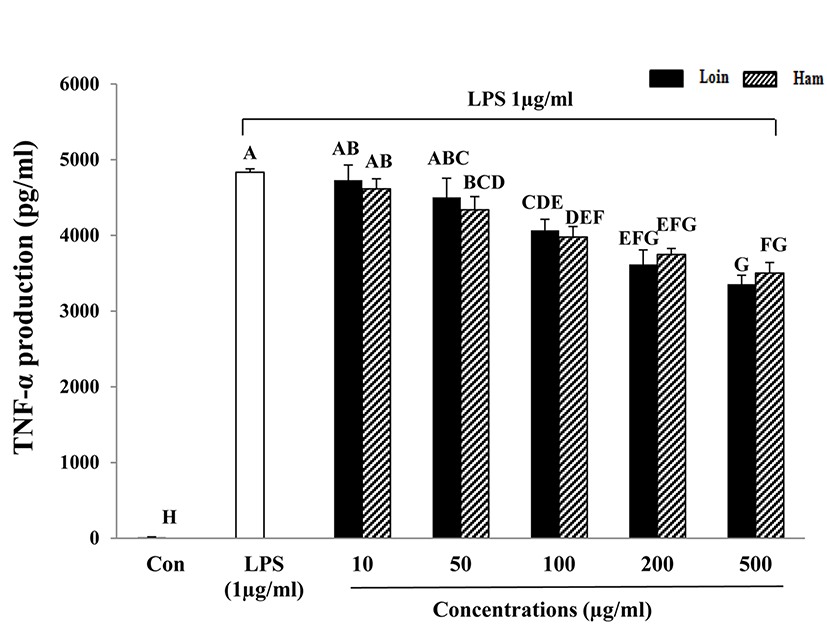
Inflammation is the response of an organism to the presence of pathogens, chemical or mechanical injury, or the first response of the immune system to infection or irritation (Hasko et al., 1996). The acute phase of inflammation in macrophages and monocytes typically plays an important role in eliciting the response cascade (Baumann and Gauldie, 1994). After they are stimulated, they generate a number of chemokines and enzymes, including cytokines, inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2) (Kim et al., 2003; Kröncke et al., 2001). LPS is a main component of the outer membrane of Gramnegative bacteria and is an endotoxin that induces septic shock syndrome by promoting the production of inflammatory mediators such as TNF-α and interleukins (Ulevitch, 1999). In addition, IL-6 is known to be a multifunctional cytokine that regulates host defense, acute phase reactions, immune responses, nerve cell functions, and hematopoiesis (Hirano, 1998). IL-6 is considered an endogenous mediator of LPS-induced fever. TNF-α secreted by activated macrophages, lymphocytes, neutrophils, and mast cells and monocytes is an inflammatory cytokine (Vileek and Lee, 1991) and is a major mediator in defense responses and the induction of apoptosis. In this study, the cytokines TNF-α and IL-6 that were secreted by LPS-activated macrophages were decreased with increased HWEP of loin and ham (Fig. 3 and 4). A dose of more than 100 μg/mL of HWEP significantly reduced the expression of IL-6 and TNF-α compared with the LPS-stimulated only group. The decrease in production of TNF-α and IL-6 appeared to attenuate cytokine-mediated host-destructive processes in inflammatory tissues. TNF-α and IL-6 secreted as inflammatory cytokines suggest a greater effect on the reduction of the inflammatory response in HWEP of loin and ham. These findings are similar to those from soy protein, which has a rich protein content similar to meat, in that they exert potent inhibitory activity toward TNF-α and IL-6 production downstream of LPS-stimulated RAW264.7 cells (Hernndez-Ledesma et al., 2009). Lee et al. (2009) provided evidence that hen egg white induced down-regulation of inflammation by reducing the expression of inflammatory cytokines such as TNF-α, IL-6, IL-1β, IFN-γ, IL-8, and IL-17. Peptides obtained from enzymatic hydrolysates of cooked tuna juice had the greatest anti-inflammatory activity in the molecular weight range of 204-1,672.9 Da (Cheng et al., 2015).
Conclusion
The HWEP of pork loin and ham has antioxidative and anti-inflammatory effects as a result of decreasing free radicals and reducing pro-inflammatory cytokines such as IL-6 and TNF-α. Further studies will be needed to determine these effects in vivo; however, this result can aid in reducing the negative perception of livestock products on human health.