










Introduction
It is currently recognized that the bacterial viability of probiotics is not an essential requirement for physiological benefits to the host (Cuevas-González et al., 2020). Accumulating evidence shows that postbiotics, that is, cell components or metabolites derived from probiotics, exert probiotic effects (Teame et al., 2020). Postbiotics are soluble bioactive compounds, such as organic acids, short-chain fatty acids, antimicrobial peptides including bacteriocins, enzymes, and vitamins (Moradi et al., 2019; Nataraj et al., 2020). Among the many beneficial effects of postbiotic compounds on the host health are antagonistic activities against pathogenic bacteria and antitumor and immunomodulatory effects (Teame et al., 2020). Several advantages of postbiotics over probiotics have been suggested, such as no risk of a blood infection, suitable absorption, directly interacting with the host and stimulating a range of biological responses, higher stability, and easier to standardize and incorporate into functional foods and pharmaceutical products (Aguilar-Toalá et al., 2018; Homayoni Rad et al., 2012).
Biofilm formed by Streptococcus mutans on tooth and dentine surfaces is a major cause of dental caries (Tong et al., 2014). During the transition from planktonic to biofilm cells, S. mutans changes its physiological characteristics (Ahn et al., 2018). For example, glucans synthesized by S. mutans from fermentable sugars, such as sucrose, via glucosyltransferases enhance bacterial adhesion to tooth surfaces and bacterial cell-to-cell adhesion, contributing to cariogenic biofilm formation (Jeong et al., 2013). It is known that bacteria in the biofilm are 10 to 1,000 times more resistant to antibiotics than planktonic bacteria (Penesyan et al., 2015). Hence, inhibiting or removing S. mutans biofilm to prevent dental caries remains a challenge in oral health care. Although a variety of cariostatic agents have been developed and clinically used to control S. mutans, their use is controversial because of some adverse side effects, including host cytotoxicity (Arabaci et al., 2013). Thus, alternative agents with minimal adverse effects are needed to control S. mutans.
Bacteriocins are small, cationic, membrane-active, antimicrobial peptides ribosomally synthesized by lactic acid bacteria with bacteriostatic or bactericidal functions (Perez et al., 2014). A most attractive feature of bacteriocins as clinical antimicrobial candidates is their GRAS (generally regarded as safe) status (Imran, 2016). A recent review has already described the role of bacteriocins in preventing biofilm formation and eradicating existing biofilms (Mathur et al., 2018). Certain antibiotic-resistant, infectious strains among enterococci, including Enterococcus faecium, have been reported, but some members of enterococci are used as probiotics (Hanchi et al., 2018). Although bacteriocin-like inhibitory substance (BLIS) produced by E. faecium DB1 exerted antimicrobial activity against Listeria monocytogenes (Choi et al., 2011), it has not been well documented whether BLIS from E. faecium DB1 (DB1 BLIS) inhibits S. mutans biofilm. Therefore, in this study, we examined whether DB1 BLIS exerts an antibiofilm effect against S. mutans.
Materials and Methods
E. faecium DB1 was kindly provided by Prof. Wang June Kim (Dongguk University, Goyang, Korea). Cariogenic S. mutans KCTC 3065 was purchased from the Korean Collection for Type Cultures (KCTC, Jeongeup, Korea). E. faecium DB1 was grown in de Man, Rogosa, and Sharpe (MRS) medium (Neogen, Lansing, MI, USA) at 37°C, and S. mutans KCTC 3065 was maintained in brain heart infusion (BHI) medium (Becton Dickinson, Sparks, MD, USA) at 37°C.
DB1 BLIS was isolated as described previously (Kim et al., 2019). In brief, E. faecium DB1 was grown in MRS broth at 37°C for 24 h. Afterward, cell-free supernatants were obtained by filtering the bacterial suspension through a 0.2-μm membrane filter. DB1 BLIS in the cell-free supernatants was precipitated by adding 70% saturation of ammonium sulfate (Samchum Chemical, Seoul, Korea) at 4°C overnight and collected by centrifugation at 17,000 g for 15 min. Precipitated DB1 BLIS was resuspended in distilled water, then placed in dialysis tubing (molecular weight cutoff 1,200; Sigma-Aldrich, St. Louis, MO, USA) and dialyzed against distilled water at 4°C for 24 h. DB1 BLIS was diluted in phosphate-buffered saline (PBS) for further experiments. The concentration of DB1 BLIS was determined by the bicinchoninic acid protein assay (Thermo Fisher Scientific, Rockford, IL, USA).
Overnight culture of S. mutans was diluted to 5 107 CFU/mL. The diluted bacterial suspension in BHI broth containing sucrose (0.05%) were transferred to wells of a 96-well microtiter plate with or without DB1 BLIS (5 mg/mL) and incubated at 37°C for the indicated times. To investigate whether the preformed biofilm of S. mutans was disrupted by DB1 BLIS, S. mutans biofilm was formed at 37°C for 24 h without DB1 BLIS. Afterward, the preformed biofilm was treated with DB1 BLIS (5 mg/mL) for 1, 3, 6, 12, and 24 h. Unattached bacteria were removed by gently rinsing with PBS and S. mutans biofilm was stained with 0.1% crystal violet for 30 min. The stained S. mutans biofilm was washed again with PBS to remove non-specific staining and dissolved in a mixture of 0.1% acetic acid and 95% ethanol. The biofilm formation by S. mutans was determined by measuring the absorbance at 595 nm using a microtiter plate reader (AMR-100, Allsheng, Hangzhou, China).
A tetrazolium salt, sodium 3’-[1-[(phenylamino)-carbonyl]-3,4-tetrazolium]-bis(4-methoxy-6-nitro)benzene-sulfonic acid hydrate (XTT), is reduced by metabolically active cells to water-soluble formazan (Wadhawan et al., 2010). Since the XTT reduction assay has commonly used for the viability of S. mutans within biofilm (Liu et al., 2017), we examined the viability of S. mutans biofilm. After biofilm formation by S. mutans in the presence or absence of DB1 BLIS (5.0 mg/mL) at 37°C for 1, 3, 6, 12, and 24 h, the biofilm was washed with PBS, followed by the addition of 100 μL of XTT (Biotium, Fermont, CA, USA) in 200 μL PBS and incubation at 37°C for 5 h. To examine the viability of preformed S. mutans biofilm cells treated with DB1 BLIS, S. mutans was incubated for 24 h, followed by DB1 BLIS (5.0 mg/mL) treatment for 1, 3, 6, 12, and 24 h. The treated S. mutans biofilm was washed with PBS, followed by the addition of 100 μL XTT in 200 μL PBS and incubation at 37°C for 5 h. The viability of S. mutans biofilm cells was determined by measuring the absorbance at 492 nm. The optical density of the background at 630 nm was subtracted from the measured absorbance at 492 nm for normalization.
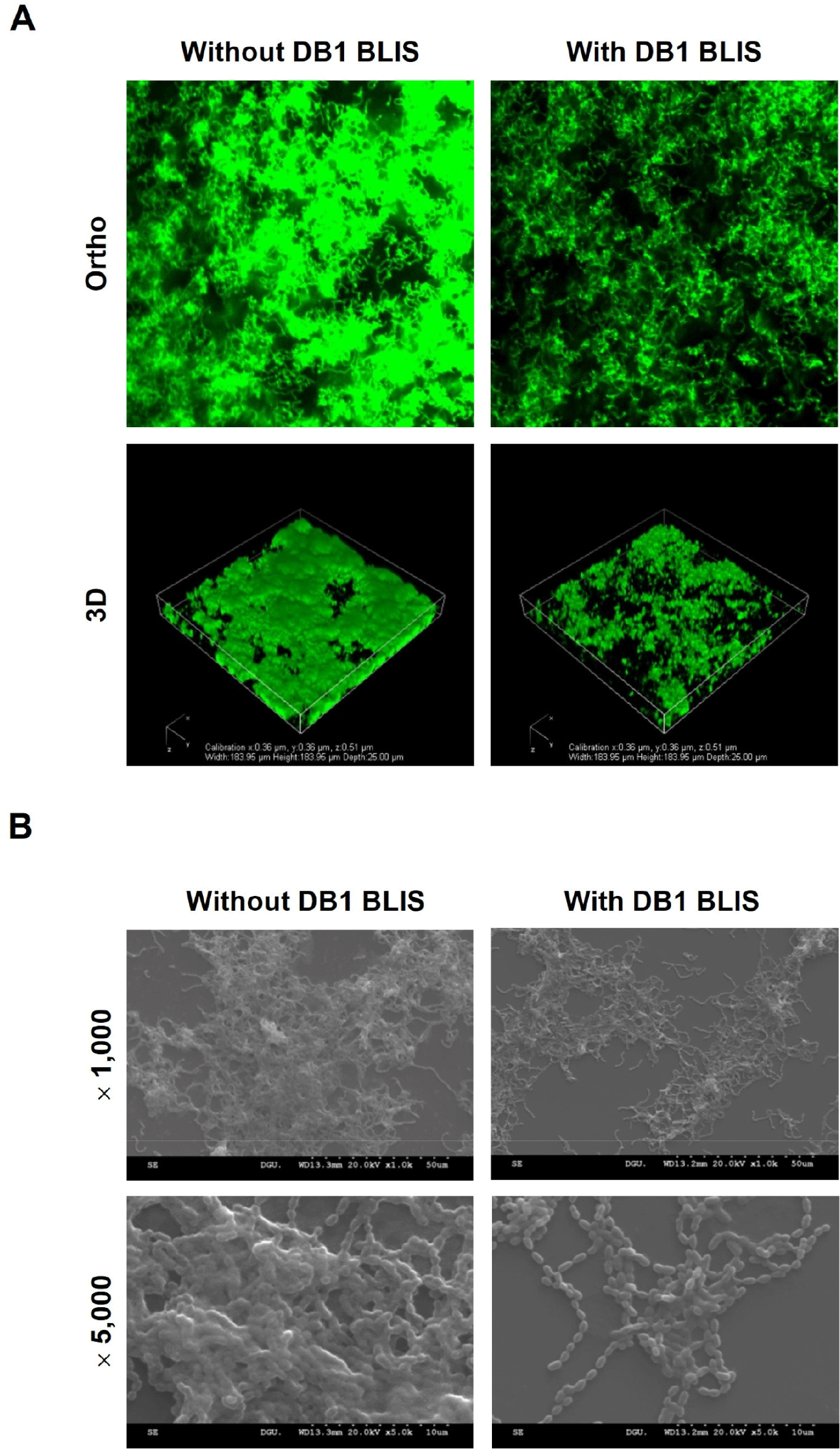
S. mutans was grown in BHI broth containing 0.05% sucrose on cover glass-bottom dishes (SPL Life Sciences, Pocheon, Korea) with or without DB1 BLIS (5.0 mg/mL) at 37°C for 24 h. After washing with PBS, S. mutans biofilm cells were stained for 10 min using the LIVE/DEAD Bacterial Viability Kit containing SYTO9 and propidium iodide (Molecular Probes, Eugene, OR, USA) according to the manufacturer’s instructions. S. mutans biofilm was observed under a confocal laser scanning microscope (Eclipse Ti-E, Nikon, Tokyo, Japan).
S. mutans was cultured in BHI broth containing 0.05% sucrose on a coverslip in the wells of a 24-well culture plate at 37°C for 24 h with or without DB1 BLIS (5.0 mg/mL). Non-adherent S. mutans was removed by washing with PBS. S. mutans biofilm was fixed with PBS containing 2.5% glutaraldehyde and 2% paraformaldehyde at 4°C overnight and dehydrated by replacing the buffer with increasing grades of ethanol (70%, 80%, 90%, 95%, and 99% for 15 min each). S. mutans biofilm was dried with hexamethyldisilazane for 15 min, sputter-coated using an ion sputter coater (E-1010, Hitachi, Tokyo, Japan), and examined under a scanning electron microscope (S-3000N, Hitachi) at 1,000 and 5,000 magnifications.
S. mutans was cultured with or without DB1 BLIS (5.0 mg/mL) in BHI broth containing 0.05% sucrose at 37°C for 1, 3, 6, 12, and 24 h. After incubation for the indicated times, the growth of S. mutans planktonic cells was determined by measuring the absorbance at 595 nm. Furthermore, the culture of S. mutans planktonic cells was serially diluted and enumerated by plating on BHI agar plates.
The saliva-coated HA assay was performed as previously described (Ansari et al., 2017). Sterile saliva-coated HA discs of 9 mm in diameter (3D Biotek, Bridgewater, NJ, USA) were placed in the wells of a 48-well microtiter plate and coated with human saliva at 37°C for 2 h. Bacterial suspension of S. mutans diluted in BHI broth containing 0.05% sucrose was added to the wells and incubated with or without DB1 BLIS at 37°C for 48 h. After incubation, saliva-coated HA discs were rinsed with PBS to remove planktonic bacteria and transferred to conical tubes containing PBS. After vigorous vortex mixing and sonication, the supernatants were collected and serially diluted for enumeration (CFU per disc) on BHI agar.
All data are expressed as mean±SD of triplicate samples. Statistical comparisons between the control and treated groups were determined at p<0.05 by unpaired two tailed t-test using GraphPad Prism 5 (GraphPad Software, La Jolla, CA, USA). Data are representative of two or three independent experiments.
Results and Discussion
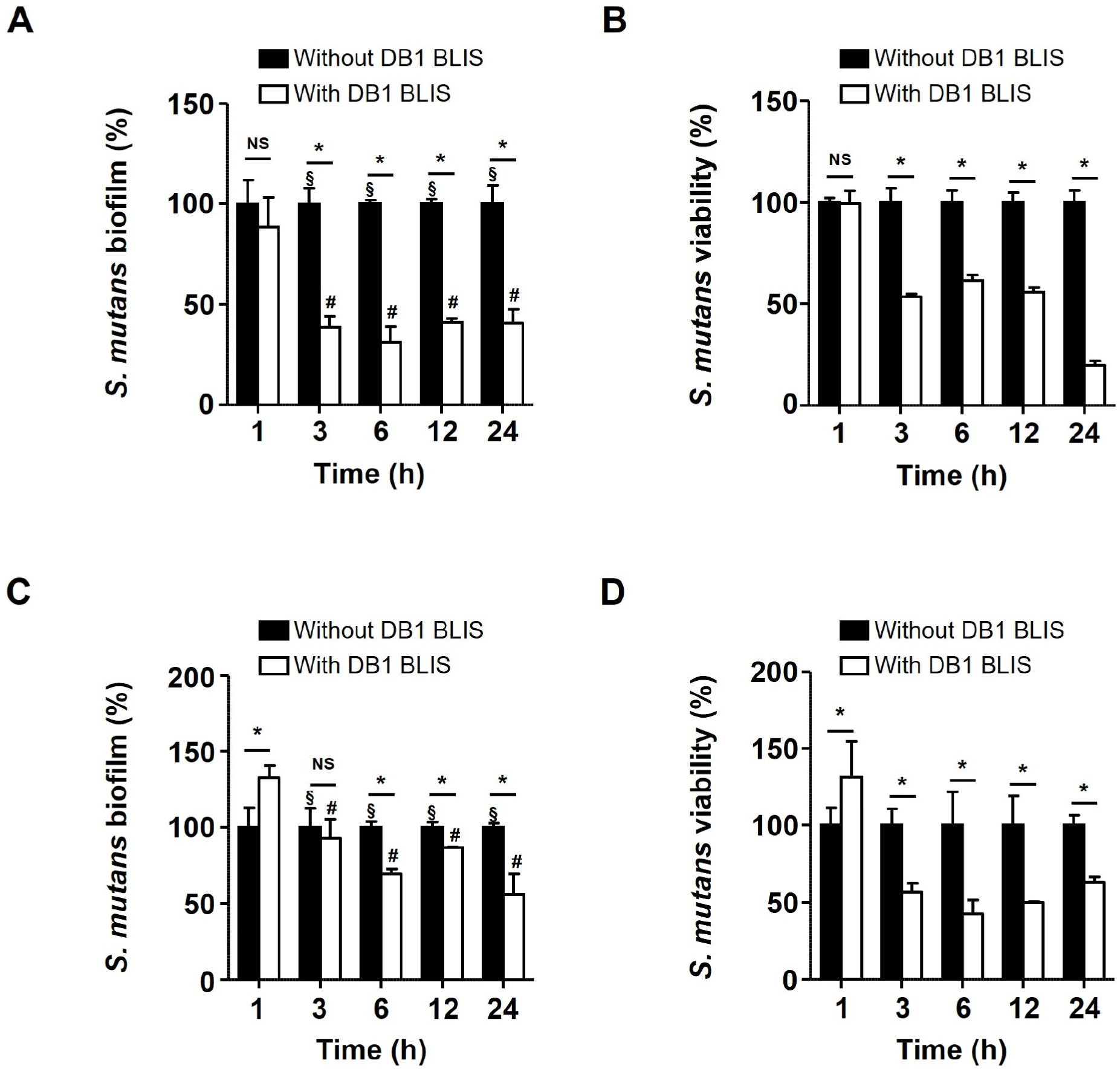
Different concentrations of DB1 BLIS (0.6, 1.2, 2.5, and 5.0 mg/mL) were tested antibiofilm activity against S. mutans. Particularly, 5.0 mg/mL of DB1 BLIS inhibited S. mutans biofilm by approximately 50% (data not shown). Therefore, this concentration of DB1 BLIS (5.0 mg/mL) was used for further experiments. When S. mutans was co-incubated with DB1 BLIS for 1, 3, 6, 12, and 24 h, DB1 BLIS significantly inhibited S. mutans biofilm at 3, 6, 12, and 24 h (p<0.05; Fig. 1A). The decreased viability of S. mutans biofilm cells in the presence of DB1 BLIS was confirmed by the XTT assay. As shown in Fig. 1B, the viability was significantly reduced at 3, 6, 12, and 24 h in the presence of DB1 BLIS (p<0.05). To examine whether DB1 BLIS eradicates established S. mutans biofilm, preformed 24-h biofilm was treated with DB1 BLIS and incubated for 1, 3, 6, 12, and 24 h. The preformed S. mutans biofilm was significantly reduced by DB1 BLIS treatment for 6, 12, and 24 h (p<0.05) but not by the shorter 1- and 3-h treatments (Fig. 1C). There was also reduced viability of preformed S. mutans biofilm cells for all treatments (3, 6, 12, and 24 h), except for the 1 h-treatment (Fig. 1D). Furthermore, microscopic analyses indicated that DB1 BLIS inhibited biofilm formation and aggregation of S. mutans. The biofilm was comprised of dense, thick layers with heterogeneously aggregated cells. However, a relatively less dense and sparsely populated biofilm was observed in the presence of DB1 BLIS (Figs. 2A and B). Studies have already shown the inhibition of S. mutans biofilm by probiotics, such as Lactobacillus plantarum (Zhang et al., 2020) and Lactobacillus salivarius (Wu et al., 2015). However, acidogenic lactobacilli are closely associated with the process of dental caries (Chhour et al., 2005; Yang et al., 2010), posing a potential dental health risk. By contrast, the bacteriocin activity of postbiotics does not have inherent cariogenic activities, such as acid production. It is widely demonstrated that bacteriocins derived from lactic acid bacteria inhibit the biofilm formation of several pathogens. Nisin of Lactococcus lactis decreased the biofilm formation of methicillin-resistant Staphylococcus aureus (Okuda et al., 2013). In our previous studies of inhibition of foodborne pathogens, we showed that Lactobacillus brevis bacteriocin effectively inhibited the biofilm formation of Escherichia coli and Salmonella Typhimurium (Kim et al., 2019), and Pediococcus acidilactici bacteriocin displayed antibiofilm activities against S. Typhimurium, Pseudomonas aeruginosa, and Enterococcus faecalis (Lee et al., 2020; Seo and Kang, 2020; Yoon and Kang, 2020). Recent studies have also shown that bacteriocins of lactic acid bacteria possess antibiofilm activity against S. mutans (Conrads et al., 2019; Dufour et al., 2020; Molham et al., 2021). In accordance with the previous reports, the current study demonstrated that DB1 BLIS could be an effective antibiofilm agent to control S. mutans.
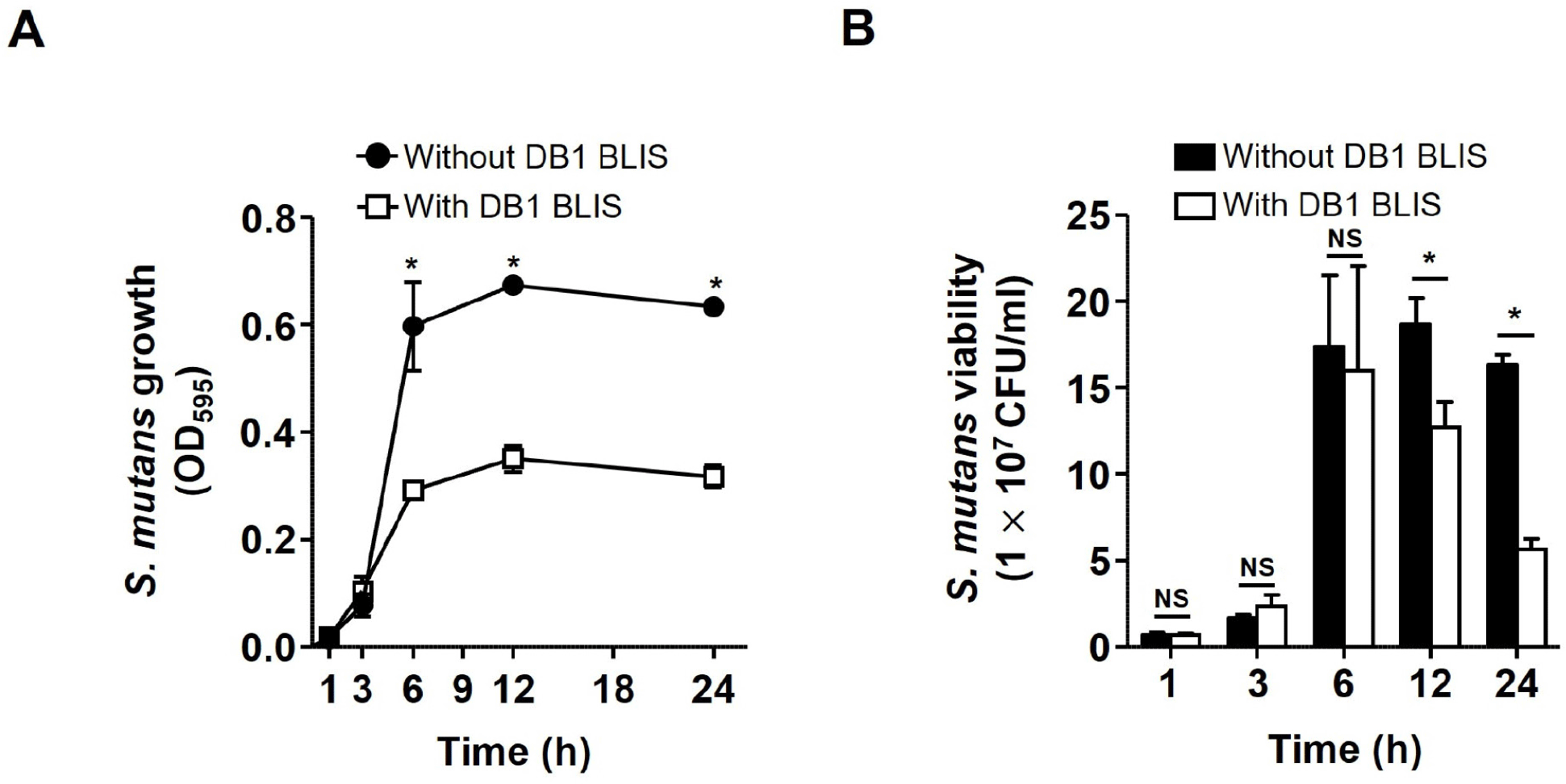
We next examined the effect of DB1 BLIS on the viability of S. mutans planktonic cells. Although short treatments with DB1 BLIS (1 and 3 h) had no effect, treatment for 6 h significantly decreased S. mutans planktonic cells viability and a decrease in viability was observed up to 24 h (p<0.05; Fig. 3A). This result was confirmed by enumeration of S. mutans planktonic cells plated on BHI agar plates. As shown in Fig. 3B, S. mutans planktonic cell count significantly decreased from approximately 1.8 108 CFU/mL without DB1 BLIS to approximately 1.3 108 CFU/mL with DB1 BLIS at 12 h (approximately 30% reduction; p<0.05), albeit that no considerable reduction in the viability of S. mutans planktonic cells was observed following shorter treatments (1, 3, and 6 h). Furthermore, DB1 BLIS more significantly reduced the viability of S. mutans planktonic cells by approximately 65%, from 1.7 108 CFU/mL without DB1 BLIS to 6 107 CFU/mL with DB1 BLIS at 24 h (p<0.05). Therefore, DB1 BLIS reduces the viability of S. mutans planktonic cells at 12 and 24 h. Since lactic acid bacteria produce different types of BLIS or bacteriocins, the antimicrobial potential of BLIS or bacteriocins may be different. In addition, bacteriocins exhibit antimicrobial potential with various spectrum depending on their types of structure, against target pathogens (Simons et al., 2020).
Bacterial attachment to the surface is the first step in biofilm. Our observations demonstrated that a decrease in the biofilm formation by S. mutans was not attributed to competitive inhibition of bacteria to the surface by DB1 BLIS. When S. Typhimurium was co-cultured with the bacteriocin of P. acidilactici HW01, both biofilm formation and viability of S. Typhimurium planktonic cells were reduced (Seo and Kang, 2020). The C-terminal 15 amino acids of synthetic human beta-defensin-3 also suppressed S. mutans growth and biofilm formation (Ahn et al., 2017). Our findings supported the previous reports that the reduction of biofilm formation by S. mutans could be associated with the inhibition of bacterial growth. Conversely, lipoteichoic acid of L. plantarum did not affect S. mutans growth but displayed exceptional ability to inhibit biofilm formation of the bacteria (Ahn et al., 2018). As mentioned above, the bacteriocin of P. acidilactici HW01 inhibited both bacterial growth and biofilm of S. Typhimurium (Seo and Kang, 2020) but markedly inhibited P. aeruginosa biofilm without suppressing bacterial growth (Lee et al., 2020). Although the inhibitory mechanism of bacteriocin or BLIS against biofilm formation remains unclear, it appears to be dependent on the target bacteria and inhibitory molecules. Most of the characterized bacteriocins of enterococci including E. faecium belong to class II bacteriocins (Nes et al., 2007). Their mechanism of antimicrobial action is based on the ability to induce membrane permeabilization, subsequently resulting in the leakage of molecules from target bacteria (Cotter et al., 2005). Our findings showed that the viability of S. mutans in biofilm was reduced by DB1 BLIS. Thus, it can be elucidated that DB1 BLIS could induce permeabilization of the cell membrane that leads to the reduction of S. mutans biofilm.
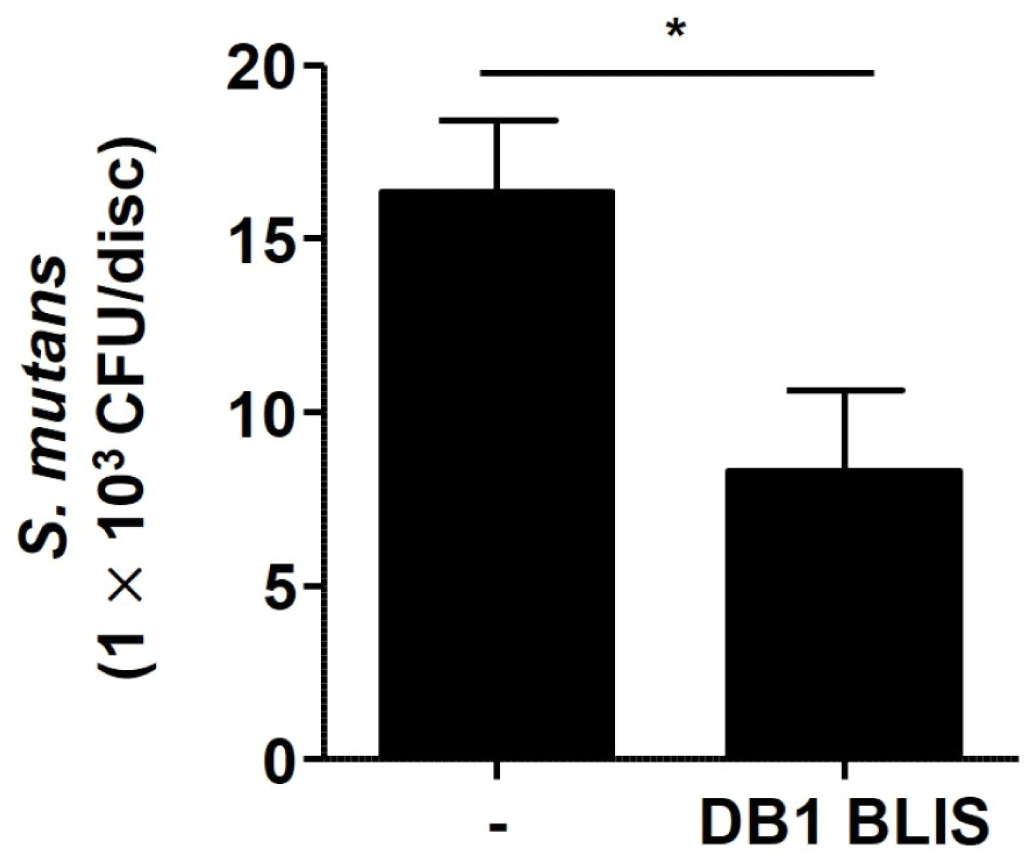
As alluded to above, inhibiting the biofilm formation of S. mutans might inhibit the occurrence of dental caries (Ahn et al., 2018). In order to examine the inhibitory effect of DB1 BLIS against S. mutans biofilm in a dental biofilm model representing dental caries, S. mutans was inoculated and allowed to form biofilm on the surface of saliva-coated HA discs with and without DB1 bacteriocin. The treatment with DB1 bacteriocin resulted in an almost 50% reduction of S. mutans biofilm (Fig. 4), suggesting the possible clinical use of DB1 bacteriocin to prevent or treat dental caries.
Besides S. mutans biofilm, multispecies bacteria in the oral cavity form biofilm, aggravating dental caries. These multispecies biofilms protect the embedded bacteria from the external environment, leading to the maintenance of biofilm architecture. Thus, it would be worth investigating the inhibitory activity of DB1 BLIS on multispecies biofilms in the oral cavity. Although further studies are needed to confirm certain molecule(s) of DB1 BLIS that are involved in the inhibition of S. mutans biofilm, our preliminary study showed that DB1 BLIS, as a postbiotic compound produced by E. faecium DB1, effectively inhibited the biofilm formation of S. mutans by decreasing the viability of S. mutans. Thus, DB1 BLIS could be a promising agent to prevent and treat dental caries caused by S. mutans biofilm.