










Introduction
The initial step of dental caries is the colonization of Streptococcus mutans, a gram-positive, facultative anaerobic bacteria, on the surface of human teeth and forming biofilms (Banas, 2004; Burne, 1998; Smith et al., 1993) because the biofilm is where other bacteria involved in dental caries attach, grow, and proliferate (Hamada and Slade, 1980). Also, the biofilm protects the bacteria from environmental stresses, antibacterial agents, and host antibodies (Stewart, 1996). The production of exopolysaccharides (EPS) and acids from carbohydrates is another important step for the cariogenic process by S. mutans (Koo et al., 2003). S. mutans use fructosyltransferase (FTF) (Shemesh and Steinberg, 2006) and glucosyltransferases (GTFs) to produce fructans and glucans from sucrose (Bowen and Koo, 2011). Fructans are extracellular storage compounds that act as binding sites for bacteria (Burne et al., 1996). Glucans play key roles in the attachment and colonization of S. mutans on the surface of the tooth, which is essential for the structural establishment of biofilms (Koo et al., 2003). Bacteria use a method called quorum-sensing to control their metabolic activity. Gene expression and communications among bacteria are important tools for them to adapt to various environmental conditions and defend themselves from the competitors and host antibodies (Shapiro, 1998). The expression of genes that regulate the production of exopolysaccharides (fructans and glucans), components of biofilm matrix, and quorum-sensing inducers in Streptococcus affects biofilm formation and dental diseases by regulating several physiological properties (Al-Sohaibani and Murugan, 2012). To prevent dental caries, therefore, both inhibiting biofilm formation by S. mutans and controlling dietary factors are important (Bradshaw and Lynch, 2013).
Most of the previous research efforts on dental caries were focused on controlling the fermentable dietary carbohydrates that serve as substrates for dental plaque (Paes Leme et al., 2006). However, foods also contain a variety of bioactive compounds that provide biological benefits (Siriwardhana et al., 2013): milk, tea, apples, and algae are considered to have beneficial effects on teeth, and some food-derived ingredients are known to inhibit caries through anti-bacterial and anti-biofilm activities (Daglia et al., 2010; Gazzani et al., 2012; Rajaraman et al., 2014; Taborsky and Mok, 1967). The egg is a well-known source of minerals, lipids, proteins, and biologically active peptides (Mine, 2007), and some of the egg proteins were found effective in inhibiting biofilm formation by preventing the adherence of various microorganisms, EPS production, demineralization, pH reduction, and biomass formation (Bradshaw and Lynch, 2013, Giacaman et al., 2019).
Phosvitin (PSV) is the major phosphoglycoprotein in egg yolk (Abe et al., 1982) with a unique structural characteristic: it contains a large number of phosphoserine groups (55% of the total amino acids) in its structure. Thus, PSV has a very strong metal-binding capability (Samaraweera et al., 2011), and shows strong antimicrobial (Lee et al., 2002) and antioxidant activities (Choi et al., 2004). The objectives of this study were to determine the effects of PSV on 1) the biofilm formation, 2) the removal of preformed biofilm, 3) the adherence ability, and 4) the expression of genes regulating the production of quorum-sensing inducers and the matrix-forming compounds (EPS) by S. mutans.
Materials and Methods
PSV was isolated from chicken egg yolk following the method of Lee et al. (2014). Crystal violet was purchased from Alfa Aesar (Haverhill, MA, USA), brain heart infusion broth (BHI) from HiMedia Laboratories (Mumbai, India), Congo red from Sigma-Aldrich (St. Louis, MO, USA), and SYBR green from Bioline (London, UK), TRIzol™ Max™ Bacterial RNA Isolation Kit from Invitrogen (Carlsbad, CA, USA), Revert Aid First-strand cDNA synthesis Kit from Thermo Fisher Scientific (Waltham, MA, USA). The S. mutans KCTC 5124, 5458, and 5316 strains were used in this study. All the strains were grown in BHI media at 37°C. The cultures were stored at –80°C in BHI containing 25% glycerol.
Biofilm formation was estimated using the crystal violet assay in a 96-well microplate (Kulshrestha et al., 2016). S. mutans were cultivated overnight in BHI media. S. mutans were diluted to 105–106 colony forming units (CFU)/mL. Fresh media (BHI+0.1% sucrose) containing 0, 0.25, 0.5, and 1 mg/mL of PSV (100 μL) and diluted S. mutans (100 μL) were added into microplate wells and the samples were incubated at 37°C under anaerobic conditions for 24 h. After incubation, planktonic cells and media were removed by gently rinsing with sterile distilled water three times. The adhered biofilm was fixed by adding 100 μL methanol for 15 min. After fixing, each well was rinsed with sterile distilled water three times and stained with 100 μL of 0.1% crystal violet for 5 min at room temperature. After staining, crystal violet was removed gently and rinsed with sterile distilled water three times. Dimethyl sulfoxide (100 μL) was added to the wells to dissolve the stained biofilm, and the absorbance was determined at 570 nm using a microplate reader (model 680, BioRad, Hercules, CA, USA).
Overnight cultured S. mutans diluted to 105–106 CFU/mL in fresh BHI media (0.1% sucrose) and 200 μL of S. mutans were inoculated to each well of a 96-well microplate. The samples were incubated at 37°C under anaerobic conditions for 24 h. The planktonic cells and media were removed and rinsed with phosphate-buffered saline (PBS). PSV at 0, 0.25, 0.5, and 1 mg/mL and media (BHI+0.1% sucrose) were added to each well and the microplate and the samples were incubated at 37°C under anaerobic conditions for 24 h. After incubation, the effects of PSV on biofilm was estimated by the crystal violet assay.
The glass surface adherence assay was used to evaluate the effects of PSV on bacterial adherence (Hamada and Slade, 1980). S. mutans 105–106 CFU/mL were transferred to glass tubes containing BHI media (0.1% sucrose) and PSV. The tubes were tilted 30 degrees and then incubated at 37°C under anaerobic conditions for 24 h. After incubation, the attached bacteria were diluted with 0.5 M sodium hydroxide and the supernatant containing planktonic cells was collected. The adhered cells and total cells were estimated by reading the absorbance at 600 nm using a spectrophotometer (Thermo Fisher Scientific). The bacterial adherence was calculated using the following formula:
Bacterial exopolysaccharide (EPS) production was evaluated by the Congo red (CR)-binding assay (Smalley et al., 1995). S. mutans 105–106 CFU/mL, media (BHI+0.1% sucrose, 200 μL) containing 0, 0.25, 0.5, and 1 mg/mL of PSV and Congo red dye (0.5 mM, 50 μL) were added to each well of a microplate, and incubated at 37°C for 1 h. At the end of incubation, the supernatant in each well was transferred to individual microtubes and centrifuged at 10,000 g for 5 min. The supernatant (200 μL) of each tube was collected and the absorbance read at 490 nm. Bacterial EPS produced was estimated using the following formula:
Quantitative real-time polymerase chain reaction (qRT-PCR) was performed to evaluate the effect of PSV on the expression of the genes that regulate the production of biofilm matrix components and quorum sensing inducers in S. mutans. Total RNA was isolated from S. mutans strains using RNA Isolation Kit and the RNA concentration was estimated at 260 nm by a spectrophotometer. cDNA was synthesized using the cDNA Synthesis Kit. The virulence gene primer sequences are shown in Table 1 (Kooltheat et al., 2016) and 16S rRNA primer used for reference gene. The SYBR green reagent was used to determine the amount of DNA. The PCR was performed for 40 cycles with a denaturing temperature at 95°C for 5 s and annealing/extension temperature at 65°C for 30 s. The qRT-PCR results were analyzed using the qRT-PCR software (PikoReal software 2.2, Thermo Fisher Scientific) that evaluates the expression of the virulence gene. The purity of PCR products was estimated using the melting curve.
Overnight cultured S. mutans (105–106 CFU/mL), media (BHI+0.1% sucrose, 200 μL) and PSV (1 mg/mL, 200 μL) were incubated at 37°C for 24 h in a 6-well microplate with glass coverslips. After incubation, the supernatants in the wells were removed and the biofilm on each glass coverslip was washed with PBS. The biofilms on the glass coverslips were fixed with 2.5% glutaraldehyde at 4°C for 1 h, washed three times with PBS and dehydrated in various concentrations of ethanol (from 50% to 100%) for 15 min. The dehydrated coverslips were dipped in isoamyl acetate for 15 min and then freeze-dried. The biofilm was scanned using a field emission scanning electron microscope (FESEM) (SU8010, Hitachi High-Technologies, Tokyo, Japan).
All results were presented as the means and the standard deviation of three replicates. The results of biofilm formation, preformed biofilm formation, EPS production, and adherence activity were performed one-way analysis of variance (ANOVA) to measure the significance of the difference in means. The student’s t-test was used to measure the significance of the difference in the means of qRT-PCR results. All results were calculated using IBM SPSS Statistics (Version 25, IBM Corp, Armonk, NY, USA).
Results
The biofilm formation and maintenance properties are an important factor of surviving in S. mutans. The inhibitory activity of PSV on the formation of biofilm by S. mutans is shown in Fig. 1. The addition of PSV at 0.25, 0.5, and 1.0 mg/mL inhibited the biofilm formation of S. mutans KCTC 5124 strain by 34.0%, 35.7%, and 42.5%, respectively, compared with the control. With the S. mutans KCTC 5458 strain, the reductions were 43.9%, 53.5%, and 58.8%, respectively, and 25.4%, 30.1%, and 33.2% with S. mutans KCTC 5316 strain. As the concentration of PSV increased, the biofilm formation by S. mutans gradually decreased, suggesting a dose-dependent anti-biofilm forming activity of PSV in S. mutans (p<0.05). The amount of preformed biofilm by S. mutans KCTC 5124 strain decreased by 25.6%, 26.9%, and 31.9% compared with the control when 0.25, 0.5, and 1.0 mg/mL of PSV was added (Fig. 2). The reduction of preformed biofilm by S. mutans KCTC 5458 strain was 21.5%, 27.5%, and 33.7% when 0.25, 0.5, and 1.0 mg/mL of PSV, respectively, was added, and that by S. mutans KCTC 5316 strain was 23.0%, 25.5%, and 33.5%, respectively. These results showed that the PSV inhibited biofilm formation and preformed biofilm of S. mtuans.
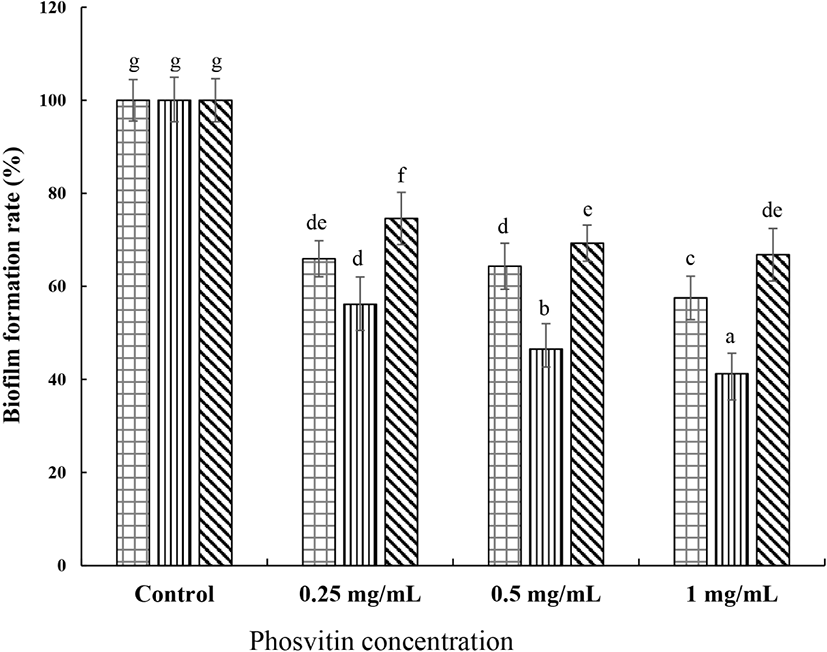
 , S. mutans KCTC 5124,
, S. mutans KCTC 5124,  , S. mutans KCTC 5458, and
, S. mutans KCTC 5458, and  , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–g Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
, S. mutans KCTC 5316. Values are expressed as the mean±SD. a–g Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
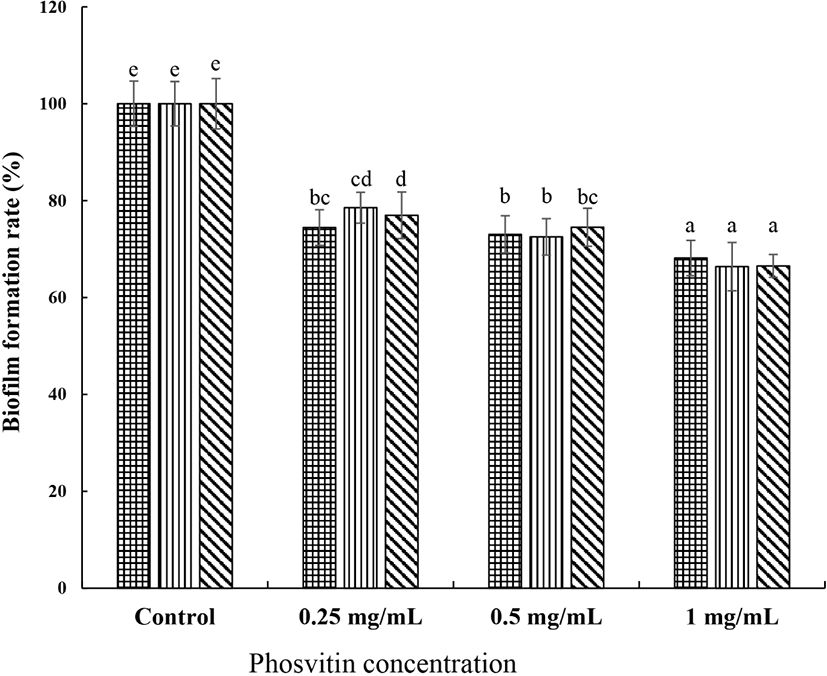 , S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–e Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
, S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–e Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
The attachment of S. mutans on the host tooth surface is first stage and key factor in colonization and biofilm formation. The effect of PSV on the adherence of S. mutans performed the glass surfaces adherence assay. Addition of 0.25, 0.5, and 1.0 mg/mL of PSV to the sample reduced 25.5%, 26.9%, and 31.9% of the adherence capacity of S. mutans KCTC 5124 strain on the glass surfaces (Fig. 3). For S. mutans KCTC 5458 strain, the reduction was 21.5%, 27.5%, and 33.6%, respectively, while that of S. mutans KCTC 5316 strain was 23.0%, 25.5%, and 33.5%. The reduction of adherence to the glass surface was dose-dependently (p<0.05). These results indicated that the PSV inhibited adherence property of S. mutans.
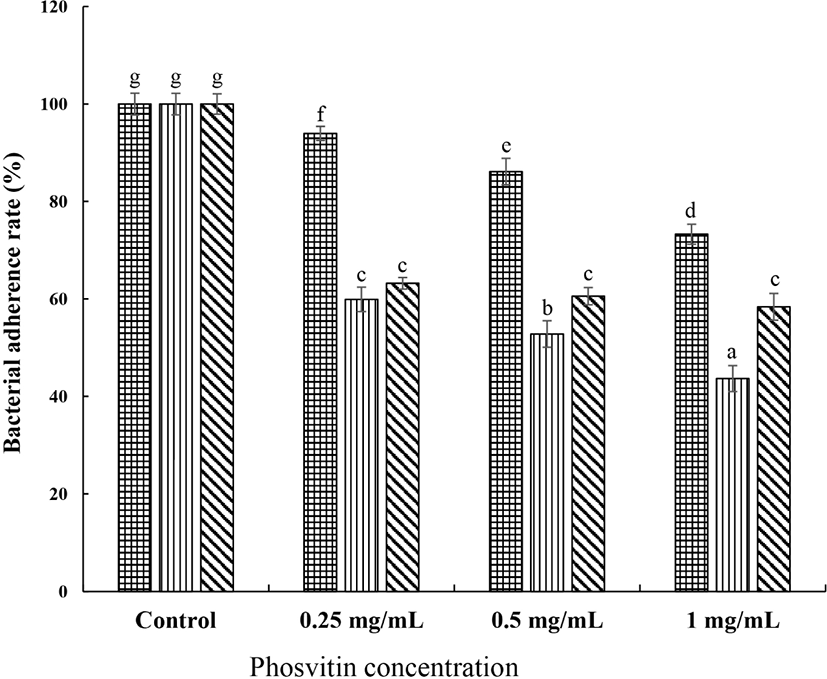 , S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–g Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
, S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–g Different letters among samples indicate significant differences (p<0.05). Control, non-treated group.
In the presence of PSV, the EPS production by S. mutans decreased dose-dependently (Fig. 4) (p<0.05). In S. mutans KCTC 5124 strain, the addition of PSV at 0.25, 0.5, and 1.0 mg/mL reduced the production of EPS by 40.8%, 51.5%, and 64.8%, respectively, compared with the control. The reductions of EPS production by the same PSV treatments to S. mutans KCTC 5458 and 5316 strain were 44.4%, 53.7%, and 65.3% and 34.0%, 43.4%, and 62.0%, respectively.
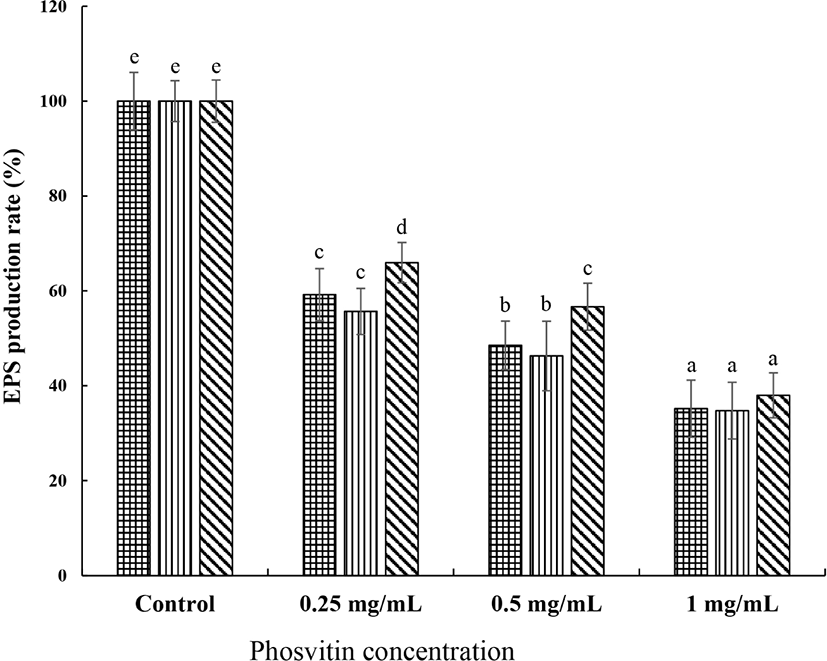 , S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–e Different letters among samples indicate significant differences (p<0.05). Control, non-treated group; EPS, extracellular polysaccharides.
, S. mutans KCTC 5124, , S. mutans KCTC 5458, and , S. mutans KCTC 5316. Values are expressed as the mean±SD. a–e Different letters among samples indicate significant differences (p<0.05). Control, non-treated group; EPS, extracellular polysaccharides.
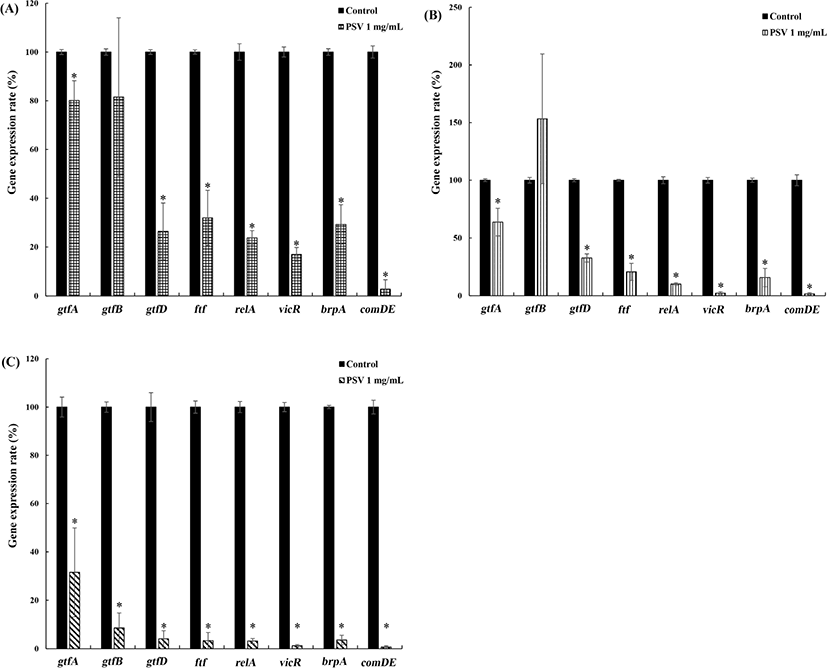
The effect of PSV on the expression of virulence genes (gtfA, gtfB, gtfD, ftf, vicR, relA, brpA, and comDE) associated with the biofilm-forming activity of S. mutans indicated that all the genes, except for gtfB, tested were significantly down-regulated by PSV (Fig. 5). The expression of gtfB decreased by 92% in KCTC 5316 strain, but there was no difference between the control group and the PSV treated group in the KCTC 5124 and 5458 strains, respectively. The expression of other virulence genes (gtfA, gtfD, ftf, relA, vicR, brpA, and comDE) was significantly down-regulated in all strains of S. mutans in the presence of PSV. However, the degree of down-regulation in KCTC 5316 strain was greater than that in other S. mutans strains.
The scanning electron microscopic (SEM) images of S. mutans strains showed dramatic reductions in biofilm formation on the glass surfaces in the presence of 1 mg/mL PSV (Fig. 6). The SEM image of S. mutans strains also showed significant reductions in the amounts of biofilms and the numbers of bacteria on the glass surfaces, which agree to the other results in this study - biofilm formation (Fig. 1), bacterial adhesion (Fig. 3) and EPS production (Fig. 4). The SEM image of control (untreated with PSV) showed a biofilm composed of multiple layers with a large number of attached bacteria, while those treated with PSV showed significantly reduced number of the attached bacteria and thin biofilm layers.

Discussion
The biofilm formation by S. mutans is the major virulence factor in developing dental caries (Hamada and Slade, 1980). The biofilm formed by S. mutans is difficult to remove because it is surrounded by a matrix composed of polysaccharides and other bacterial biofilms. The key to the inhibition of biofilm is suppressing its formation at an early stage (Islam et al., 2008). PSV showed dose-dependent activity in the reduction of the preformed biofilm by S. mutans (p<0.05). PSV significantly inhibited the formation of biofilm by S. mutans at an early stage. Also, the amounts of preformed biofilms by S. mutans decreased when PSV was present, suggesting that PSV not only prevented the formation of biofilms but also degraded the preformed biofilms by S. mutans. Thus, PSV has great potential to be used to prevent dental caries.
In general, bacteria are preferentially colonized on the surfaces that are hydrophobic and with roughness surfaces (Bowen and Koo, 2011). S. mutans starts to colonize on the teeth surface after adhesion using sucrose-dependent and sucrose-independent processes. The sucrose-dependent attachment process is associated with the production of glucans and fructans, while the sucrose-independent attachment process is associated with the electrostatic forces, cell-surface-binding proteins, and hydrophobic interactions (Staat et al., 1980). PSV inhibited the adhesion of S. mutans cells on the glass surfaces by changing the biofilm matrix. PSV inhibited the sucrose-dependent attachment by down-regulating carbohydrate metabolism and the sucrose-dependent EPS production. Also, PSV inhibited the initial sucrose-independent attachment by down-regulated comDE gene and inhibited LytR. The decrease of adhesion is expected to have a marked inhibitory effect on early colonization and biofilm formation by S. mutans.
Polysaccharides are the major part of the biofilm matrix and the exopolysaccharides produced by S. mutans are the key factor to form indispensable mature biofilms, the maintenance, and expansion of the biofilms (Flemming and Wingender, 2010). Thus, the amounts and types of exopolysaccharides produced are very important for the formation, maintenance of the bacterial community, and the structure of biofilms (Sutherland, 2001). This indicated that the significant reduction of EPS production by S. mutans in the presence of PSV is very important for the anti-biofilm-forming ability of PSV.
The downregulation of virulence genes in S. mutans by PSV indicated that the inhibitory activity of PSV in the biofilm formation by S. mutans is related to its effect on gene expression. The gtfA genes encode GTF-A enzyme, a sucrose phosphorylase (Russell et al., 1988). The gtfB encodes water-insoluble glucans, the adhesion molecules that are essential for the sucrose-dependent attachment and immobilization of bacteria on hard surfaces (Wen et al., 2010). The gtfD produces water-soluble glucans that are sucrose-dependent (Hanada and Kuramitsu, 1988). Although, in KCTC 5124 and 5458 strains PSV did not affect to expression of gtfB, gene expression of gtfA and gtfD was decreased, and similarly, EPS formation was also decreased. Therefore, PSV suppressed the expression and activity of GTFs, which lowered the formation of EPS. These results indicated that PSV has anti-biofilm activity by inhibiting the production of extracellular polysaccharides (EPS), the key components of the biofilm matrix. The brpA encodes the surface-associated biofilm-regulatory protein that is vital to biofilm formation and the responses to the environmental stresses (Wen and Burne, 2004). The LytR-CpsA-Psr family proteins encoded by brpA. LytR has an important role in the cell division and in sucrose-independent attachment (Chatfield et al., 2005; Yoshida and Kuramitsu, 2002). The decreased expression of brpA gene by PSV compromised the stress resistance of S. mutans. The vicR gene encodes a two-component regulatory system that regulates the expression of several genes related to the sucrose-dependent adherence and the synthesis of polysaccharides, including ftf and gtf gene family (Senadheera et al., 2005). The vicR gene is down-regulated by PSV and the suppression of this gene led to an anti-biofilm forming effect through the inhibition of adherence activity. Various factors, which include environmental stresses and intracellular processes such as carbohydrate metabolism can influence the expression of GTFs (Banas and Vickerman, 2003). Quorum-sensing is an important mechanism in the formation of biofilms by S. mutans. The relA and comDE genes contribute to the quorum-sensing and biofilm formation by S. mutans. The relA gene encodes RelA, a carbohydrate phosphotransferase system (PTS) protein that regulates the glucose uptake system by phosphoenolpyruvate (Lemos et al., 2004). The comDE gene encodes two-component signal transduction systems composed of comD and comE that are part of competence-stimulating peptides. The comD and comE act as histidine kinase receptors and cognate response-regulators in the quorum-sensing of S. mutans (Suntharalingam and Cvitkovitch, 2005). If comD and comE are defective, biofilms with reduced biomass can be formed and initial sucrose-independent attachment (Li et al., 2002). The expression of relA and comDE were also down-regulated, especially the expression of comDE, by PSV and the decrease was over 90% in all strains. These results indicated that PSV inhibited the biofilm-forming activity of S. mutans by controlling the regulatory genes involved in the quorum-sensing system and the production of the key components for biofilm matrix formation.
Conclusion
It can be concluded that PSV showed cariostatic properties through the following mechanisms: 1) inhibiting biofilm formation by S. mutans and the degradation of pre-formed biofilms, 2) inhibiting the production of EPS, and 3) inhibiting bacterial adhesion to the surfaces, and all these inhibiting activities of PSV are through the control of gene expression in S. mutans. PSV down-regulated the expression of genes related to the production of EPS, autoinducers of quorum sensing, and the key components of the biofilm matrix. Thus, PSV has high potentials to be used as a treatment as well as a preventive agent for dental caries.