










Introduction
Staphylococcus aureus is one of the most notorious pathogenic bacteria and resides in various natural environments, including human skin (Feuerstein et al., 2017). In the dairy industry, the pathogenic bacteria are seriously controlled microbes because they can cause bovine mastitis and easily can enter the dairy chain (Kummel et al., 2016). Therefore, it is very important to effectively control S. aureus in the dairy industry. Besides the industrial perspective, multi-drug resistant S. aureus is a serious threat in clinical areas and alternative control agents are needed (Hiramatsu et al., 2014).
Bacteriocins from lactic acid bacteria (LAB) are powerful antimicrobial peptides that strongly inhibit the growth of pathogenic bacteria, such as S. aureus and Listeria monocytogenes (Juturu and Wu, 2018). To date, many bacteriocins from LAB have been purified and identified (Jamaluddin et al., 2018). Specifically, nisin from Lactococcus lactis was commercialized and is widely used in foodstuffs as a biopreservative (Gough et al., 2017).
A bacteriophage is a virus that infects target bacteria. Due to the emergence of antibiotic-resistant pathogenic bacteria, phage therapy is strongly considered as an alternative antibiotic therapy. Unlike antibiotics, bacteriophages have the benefit of species-specificity, meaning that only the target bacterial species can be controlled by the bacteriophage, without disturbing natural microflora harboring beneficial or desirable bacteria (Kakasis and Panitsa, 2019).
Previously, a Lactococcus lactis strain that produces a bacteriocin showing strong antimicrobial activity against S. aureus was isolated and a bacteriophage that specifically infects S. aureus was identified. In this study, we investigated the synergistic inhibition of S. aureus by the bacteriocin and the bacteriophage combination.
Materials and Methods
To prepare crude bacteriocin, a modified acetone extraction method was used (Chung et al., 2011). Briefly, Lactococcus lactis CJNU 3001 (GenBank accession number: MN749817), which is the bacteriocin producer and isolated from kimchi, was inoculated into 100 mL of MRS (de Man, Rogosa, and Sharpe) broth and cultured at 30°C for 12 h. The culture was centrifuged at 6,500×g for 10 min at 4°C to recover the culture supernatant and filtered with a 0.45-μm membrane filter (Agela Technologies Inc., Wilmington, DE, USA) to eliminate the bacterial cells. The filtrate was concentrated with a rotary evaporator (Eyela, Tokyo, Japan) and mixed with acetone (concentrate:acetone=1:3, v/v). The mixture was stored at −20°C for 3 h and vigorously shaken every 30 min. It was centrifuged at 8,000×g for 20 min at 4°C and the middle layer was recovered and filtered through the membrane filter. The crude bacteriocin was serially diluted (2-fold) and 2 μL of each diluent was loaded on an MRS agar plate where S. aureus KCTC 3881 had been seeded to measure the bacteriocin activity (arbitrary units, AU; Daeschel, 1992).
To propagate the bacteriophage SAP84, which is specific to S. aureus and isolated from soil, S. aureus KCTC 3881 and the bacteriophage SAP84 were inoculated into 5 mL of MRS broth at a ratio of 2.5:1 and cultured at 37°C for 6 h with shaking. The culture was centrifuged at 6,500×g for 10 min at 4°C and the culture supernatant was filtered with the membrane filter. The filtrate was serially diluted (10-fold) and 10 μL of adequate diluent was loaded on an MRS agar plate where S. aureus KCTC 3881 had been seeded to count the number of plaques (plaque-forming units, PFU).
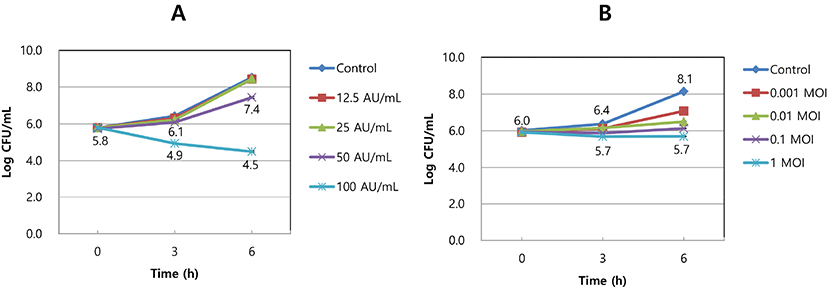
Approximately 1.0×106 CFU/mL of S. aureus KCTC 3881 was inoculated into 5 mL of MRS broth. Bacteriocin at 12.5, 25, 50, or 100 AU/mL was added to the broth and incubated at 37°C for 6 h. The viable cell count of the strain was measured at different times, and the results are presented as CFU/mL.
Approximately 1.0×106 CFU/mL of S. aureus KCTC 3881 was inoculated into 5 mL of MRS broth. The SAP84 bacteriophage at 0.001, 0.01, 0.1 or 1 MOI (multiplicity of infection) was added to the broth and incubated at 37°C for 6 h. The viable cells of the strain were counted at different times and are presented as CFU/mL.
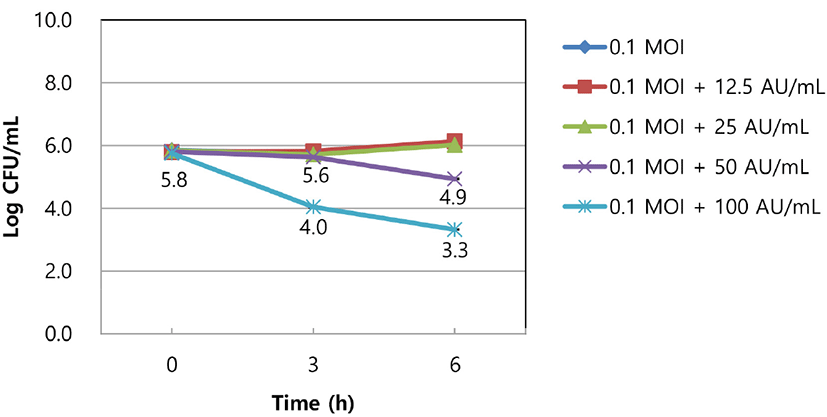
Approximately 1.0×106 CFU/mL of S. aureus KCTC 3881 was inoculated into 5 mL of MRS broth and 0.1 MOI of the SAP84 bacteriophage was added to the broth. Bacteriocin at 12.5, 25, 50, or 100 AU/mL was added to the broth and incubated at 37°C for 6 h. The viable cells of the strain were counted at different times and are presented as CFU/mL.
Results and Discussion
Bacteriocin from L. lactis CJNU 3001 was partially purified by the modified acetone extraction method and the crude bacteriocin activity against S. aureus KCTC 3881 was shown to be 32,000 AU/mL (Fig. 1A). To propagate the bacteriophage SAP84, S. aureus KCTC 3881 was used as a host strain. After 6 h incubation with the strain, the bacteriophage SAP84 reached 1.3×109 PFU/mL (Fig. 1B).

The crude bacteriocin showed antimicrobial activity against S. aureus KCTC 3881 in a dose-dependent manner. Bacteriocin activity units of 12.5 and 25 AU/mL did not affect the growth of S. aureus KCTC 3881, but 50 and 100 AU/mL inhibited the growth of the strain proportionally to the dose. Specifically, a nearly 4-Log CFU/mL reduction in the strain was seen after 100 AU/mL of bacteriocin treatment and 6 h of incubation compared to the control (no addition of the bacteriocin; Fig. 2A). In the experiment of SAP84 bacteriophage, after 6 h of incubation, the viable cell count of S. aureus KCTC 3881 reached 8.1 Log CFU/mL. However, the cell count was reduced when cultured with the bacteriophage SAP84 in a dose-dependent manner. When the strain in MRS broth was treated with 1 MOI of SAP84 bacteriophage, the viable cell count reached 5.7 Log CFU/mL after 6 h of incubation (Fig. 2B).
To investigate the synergistic inhibition of the bacteriocin and the bacteriophage SAP84 on the growth of S. aureus KCTC 3881, 0.1 MOI of the bacteriophage concentration was combined with various bacteriocin activity units ranging from 12.5 to 100 AU/mL. The viable cell counts of the strain were not changed by treatment with 12.5 or 25 AU/mL bacteriocin combined with 0.1 MOI of the bacteriophage. However, the phage showed synergistic inhibition against the strain when combined with 50 or 100 AU/mL of the bacteriocin. The viable cell counts of both treatments reached 4.9 and 3.3 Log CFU/mL after 6 h of incubation, whereas the control (only the bacteriophage was treated) reached 6.1 Log CFU/mL (Fig. 3).
The use of bacteriocin or bacteriophage for controlling pathogenic bacteria including S. aureus has been widely studied, but the combination of both antimicrobials for the same purpose has been rarely studied. Yuksel et al. (2018) investigated the synergistic inhibition by the combinations of bacteriophage, EDTA, and nisin on the formation of Salmonella Typhimurium biofilm. In the study, the combination of nisin and EDTA showed a synergistic effect but the combination of nisin and bacteriophage did not. Hathaway et al. (2017) designed a nanoparticle harboring a bacteriophage endolysin and a bacteriocin for controlling methicillin-resistant S. aureus (MRSA) on human skin that could release the included molecules by temperature control. The nanoparticles worked effectively at 37°C, the temperature associated with skin wounded by infection, and cell lysis was observed. However, at 32°C, which is normal skin temperature, they did not work. Garcia et al. (2010) reported a synergistic effect of the bacteriophage endolysin LysH5 and nisin to control S. aureus in milk. In the study, only the combination of both antimicrobials cleared the viable cells of S. aureus from milk after 6 h of incubation at 37°C. Additionally, the authors knew that the activity of the LysH5 endolysin increases in the presence of CaCl2, MgCl2, and NaCl, whereas it decreases in the presence of MnCl2 and ZnCl2. Therefore, they optimized the buffer conditions for the best activity of the endolysin LysH5 as 50 mM phosphate buffer, 1 mM CaCl2, 1 mM MgCl2, and 100 mM NaCl at pH 7.0. Previously, we also tried to test the synergistic effect of an acidic bacteriocin, like nisin, and a purified endolysin from a bacteriophage targeting S. aureus. However, a synergistic effect due to the different optimal pHs for the activities of the two different antimicrobials was not shown (unpublished data). Furthermore, purification from recombinant Escherichia coli cell lysate is necessary to obtain purified endolysins, which is time-consuming and not cost-effective. Therefore, a direct use of the bacteriophage with the bacteriocin to present a synergistic inhibition against pathogenic S. aureus was determined. In this study, we showed synergistic inhibition by the bacteriophage SAP84 and the bacteriocin from L. lactis CJNU 3001 even though an efficacy of the combination were not proved in real food system since antimicrobial activity of bacteriocin or bacteriophage might be influenced by food matrix effects. Based on the results, future studies to optimize the activity of the combination and test its efficacy in the industrial field will be conducted.