










Introduction
Lactic acid bacteria (LAB) represent probiotic bacteria (Sarao and Arora, 2017) and have demonstrated positive physiological effects on human health that include controlling pathogens, improving constipation, alleviating diarrhea, lowering blood cholesterol, as well as immunological activity, and anti-tumor effects, etc. (Elbanna et al., 2018; Juturu and Wu, 2018; Liu et al., 2017; Mallina et al., 2018; Shida and Nomoto, 2013; Wojtyniak and Szajewska, 2017). Recently, the bacteria have received more attention since they comprise the main commensal bacteria in our gut where various phyla co-exist and communicate with host cells (Azad et al., 2018; Qin et al., 2010).
Bacteriocins are ribosomally synthesized antimicrobial peptides which inhibit closely related species of the producers. In particular, bacteriocins from lactic acid bacteria have been widely studied for safety reasons and nisin, produced by Lactococcus lactis, is commercialized and used in foods as a biopreservative (Dicks et al., 2018; Jutura and Wu, 2018).
Staphylococcus aureus is pathogenic bacteria that resides in various environments, including the human body, as well as foods regulated by safety authorities and organizations (Fetsch et al., 2014; Fitzgerald and Holden, 2016; Muenks et al., 2016). In clinical areas such as hospitals, the bacteria are intensively treated and antibiotic resistant strains, such as MRSA (methicillin resistant S. aureus) and VRSA (vancomycin resistant S. aureus), are frequently detected (Gould 2013; McGuinness et al., 2017). Therefore, it is necessary to develop alternative antimicrobial agents instead of antibiotics to control the pathogenic bacteria in food or food materials.
In this study, we screened lactic acid bacteria producing bacteriocin which inhibited the growth of S. aureus and investigated its preliminary characteristics.
Materials and Methods
LAB and S. aureus strains were cultured in MRS (De Man, Rogosa, and Sharpe) broth (Difco, Sparks, MD, USA) at 37℃ without shaking.
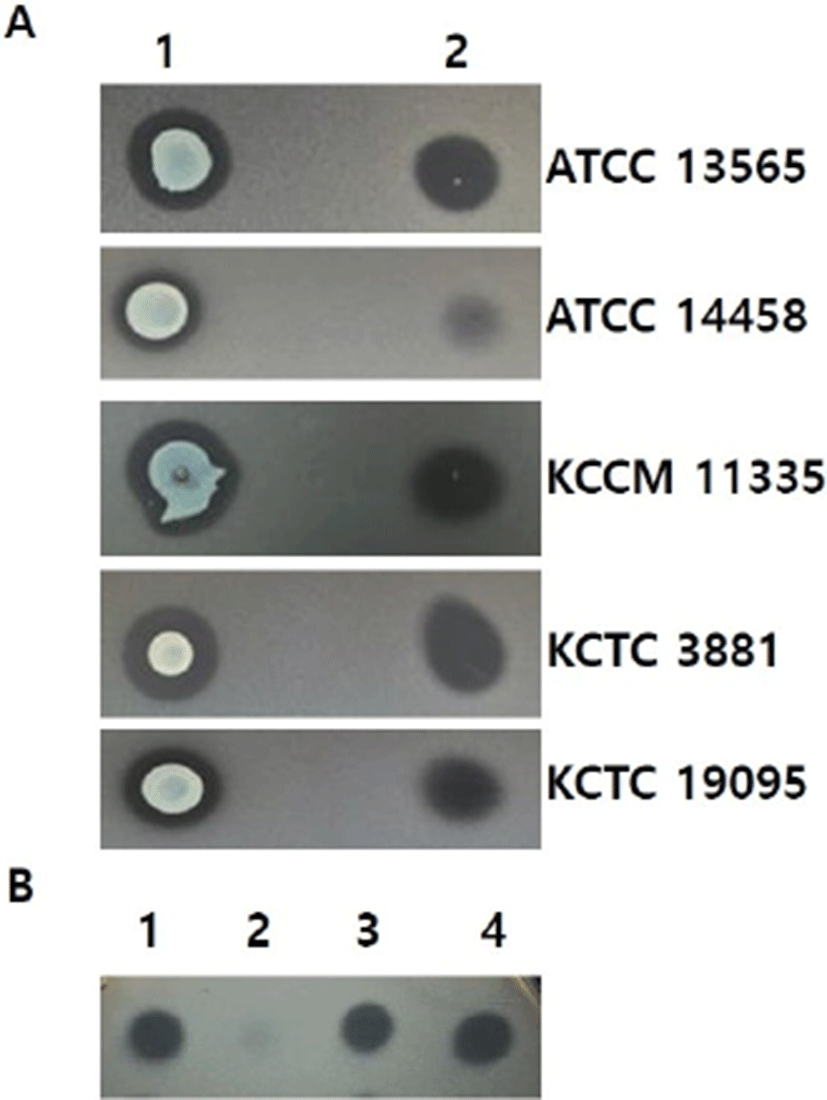
Almost 1,000 LAB strains from kimchi were screened for production of anti-staphylococcal bacteriocin. The LAB cultures were spotted on MRS agar plates and incubated for 6 h and S. aureus indicator strains (ATCC 13565, ATCC 14458, KCCM 11335, KCTC 3881, and KCTC 19095) were overlaid on the plates with MRS soft agar (0.7% agar). The plates were incubated for another 12 h and inhibition zones around colonies were examined (Moon et al., 2000; Daeschel, 1992). The strain showing anti-staphylococcal bacteriocin activity was identified by 16S rRNA gene sequencing performed by a commercial biotech company (Macrogen, Daejeon, Korea) and homology search by using a BLAST program (blastn) served from NCBI (http://www.ncbi.nlm.nih.gov/).
The strain presenting anti-staphylococcal bacteriocin activity was cultured in several environments: i) MRS or RCM broth as a culture medium and ii) 25℃, 30℃, or 37℃ as incubation temperatures. During the incubation for 24 h, samples were taken and viable cell counts (CFU/mL) and bacteriocin activities (AU/mL) were measured at 0, 3, 6, 9, 12, and 24 h. The AU (arbitrary units) denote the reciprocal of the highest two-fold dilution showing an inhibition zone (Daeschel, 1992).
The bacteriocin was partially purified by acetone extraction (Chung et al., 2011) and it was tested for resistance against heat treatments (60℃–121℃), organic solvents such as methanol (Honeywell B & J, Ulsan, Korea), ethanol (Honeywell B & J), acetone (Honeywell B & J), acetonitrile (Honeywell B & J), and chloroform (Avantor Performance Materials, Center Valley, PA, USA), and enzymes such as protease (Tokyo chemical, Tokyo, Japan), lipase (Tokyo chemical), and α-amylase (Sigma-Aldrich, St. Louis, USA).
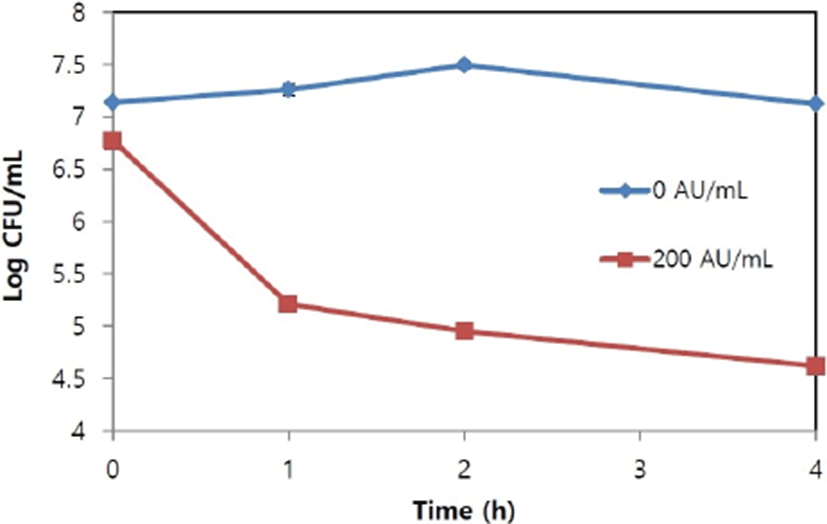
The culture of S. aureus KCTC 3881 strain was added into 30 mL of peptone water (0.1%, w/v), 200 AU/mL of the bacteriocin was added to the inoculum and viable cell counts (CFU/mL) were measured at 0, 1, 2, 3, and 4 h. Based on the measurements, we concluded that the bacteriocin showed either bacteriostatic activity (growth inhibition, but no killing) or bactericidal activity (bacteria killed).
Results and Discussion
Among 1,000 LAB strains, only one strain, CJNU 3001, showed antimicrobial activity against S. aureus strains (ATCC 13565, ATCC 14458, KCCM 11335, KCTC 3881, and KCTC 19095). In particular, the bacteriocin from CJNU 3001 strain showed the strongest antimicrobial activity against S. aureus KCTC 3881 and we chose that strain as the indicator strain for subsequent experiments (Fig. 1A). The 16S rRNA gene sequence of CJNU 3001 strain was analyzed by a commercial company (Macrogen) and aligned by using a BLAST program (blastn) from NCBI for homology search. The results showed that the sequence of CJNU 3001 was identical to Lactococcus lactis (100% identity; accession no. NR_040955.1) and the strain named L. lactis CJNU 3001.
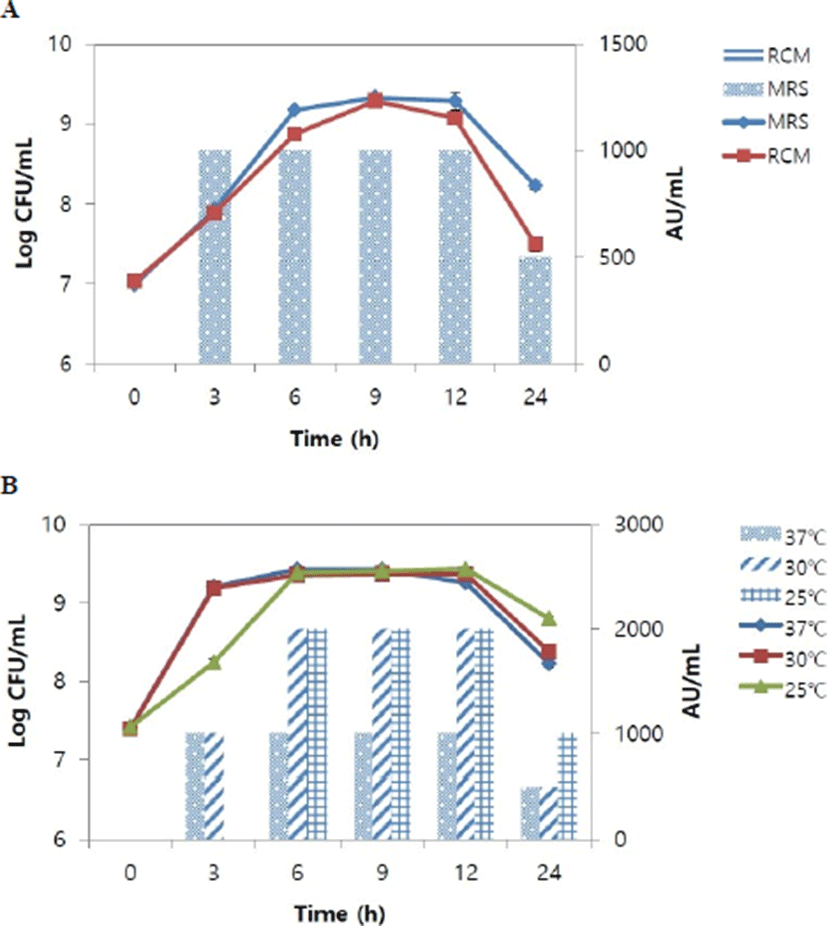
To select the optimal medium for production of bacteriocin, L. lactis CJNU 3001 was incubated in MRS or RCM broth. The viable cell counts in both media were not significantly different with each other (Fig. 2A). The counts reached the maximum level at 9 h where 9.3 Log CFU/mL for both MRS broth and RCM broth was measured. However, bacteriocin activity was detected only in MRS broth, at a maximum level of 1,000 AU/mL that was maintained for 3–12 h (Fig. 2A). Using MRS broth, L. lactis CJNU 3001 inocula were incubated at 25℃, 30℃, or 37℃ to select the optimal temperature for production of the bacteriocin. The viable cell counts at different culture conditions were not significantly different except for 3 h where the count (8.2 Log CFU/mL) at 25℃ was even lower than those (9.2 Log CFU/mL) of 30 and 37℃ (Fig. 2B). The bacteriocin activity was the highest at 25℃ and 30℃, measured at 2,000 AU/mL that lasted 6–12 h, whereas 1,000 AU/mL activity seen at 37℃ lasted 3–12 h. The bacteriocin activity at 25℃ was maintained at 1,000 AU/mL until 24 h but the activity decreased to 500 AU/mL at 30℃ and 37℃ (Fig. 2B). Therefore, the optimal conditions for the production of the bacteriocin from L. lactis CJNU 3001 were MRS broth incubated at 25℃.
The partially purified bacteriocin was stable at 60℃ or 80℃ for 60 min, and 15 min at 100℃, compared to the control production of 4,000 AU/mL that decreased to 2,000 AU/mL at 100℃ at 30 min and 500 AU/mL 60 min, and 500 AU/mL at 121℃ for 15 min (Table 1). The bacteriocin was also stable under treatment of organic solvents such as methanol, ethanol, acetone, acetonitrile, and chloroform (Table 1). It was resistant to enzymes lipase and α-amylase, but not to protease which further supports that the antimicrobial agent of L. lactis CJNU 3001 is a bacteriocin (Fig. 1B).
The bacteriocin showed bactericidal action indicating it killed the target bacteria S. aureus KCTC 3881. The viable cell count of S. aureus KCTC 3881 started 7.2 Log CFU/mL, but decreased to 4.6 Log CFU/mL 4 h after 200 AU/mL of bacteriocin was added, whereas it started at 7.2 Log CFU/mL and reached 7.1 Log CFU/mL after 4 h incubation in the control where no bacteriocin was added (Fig. 3).
As mentioned above, S. aureus is one of notorious human pathogenic bacteria and widely contaminates food materials including meats and vegetables (Paudyal et al., 2017). In particular, MRSA and VRSA which originated from clinical areas like hospitals due to wide-spread antibiotic use are threatening our health (Kos et al., 2012). To alleviate this threat, alternative antimicrobials need to be identified to control the pathogen. Bacteriocins from LAB are safe for human use and have been broadly studied (Etayash et al., 2015). They are recognized as one of the next generation antibiotics and can be commercialized (Chikindas et al., 2018; Seal et al., 2018). The bacteriocin from L. lactis CJNU 3001 showed antimicrobial activity and bactericidal action against several S. aureus strains. It was stable at heat treatments and solvents treatments which are good characteristics for applying to industrial fields. Even though the results in this study are preliminary, the bacteriocin might represent a promising new antimicrobial agent or a food biopreservative to control the pathogen. In near future, we will purify the bacteriocin and biochemical and genetic traits of the bacteriocin will be characterized for mass production for commercialization.