










Introduction
Infant formula (IF) is defined as an infant diet in a powdered form that mimics human breast milk (HBM), and therefore can be used as a substitute of HBM (FDA, 2019). HBM consists of 87% water, 3.8% fat, 1.0% protein, and 7% lactose (Martin et al., 2016). IF is usually prepared using a bovine milk base, which is known to have higher protein and lower carbohydrate levels than HBM (Wargo, 2016). Carbohydrates are the major source of energy for humans when converted in glucose by intestinal digestion. Carbohydrates are important because of their necessity in childhood growth (Kalhan and Kilic, 1999). These compositional differences hamper infant growth and lead to the development of certain childhood illnesses such as sore throat, ear pain and skin infections (Boyd et al., 2007). Boué et al. (2018) suggested that HBM oligosaccharides might lower the risk of developing gastrointestinal (GI) infections and respiratory tract infections by inhibiting the adherence of pathogens to the gut mucosa. Neurodevelopment is also enhanced by HBM feeding, possibly by DHA transmission, contributing to structural changes in the brain through FADS2 gene regulation (Horta and Victora, 2013). Fortification of HBM using IF, improved growth parameters such as weight gain and head growth in low-birth-weight infants (Gupta et al., 2020).
To narrow the compositional and functional gap between HBM and IF, IF fortification has been implemented by supplementation with additives such as micronutrients, prebiotics, and probiotics. Consumption of iron-containing fortified milk for a year reduced anemia and iron deficiency in 12-to 30-month-old children (Brown Kenneth et al., 2007). Consumption of IF supplemented with prebiotics, galactooligosaccharides, and fructooligosaccharides, increased bifidobacterial colonization in the infant GI tract (Borewicz et al., 2019). Bifidobacterium is a predominant bacterium found in HBM-fed infants and confers protection to the host against pathogens by enhancing mucosal immune system functions (Makino, 2018). Probiotic-enriched IF, containing Bifidobacterium animalis, altered the gut microbiota (GM) pattern, fecal IgA content, and stool pH to resemble those in HBM-fed infants (Radke et al., 2017).
The GM, which is a complex microbial community of bacteria, archaea, fungi, viruses, and phages living in the gut, reportedly controls various host pathways (Derrien et al., 2019). Infant GM regulates infant metabolism, nutrient absorption, immune system maturation, and pathogen colonization (Belkaid and Hand, 2014). The development of GM is shaped by exterior factors such as diet, environment, and lifestyle; therefore, there exists a unique gut microbial composition specific to each individual (Derrien et al., 2019).
Infant diets contribute to the formation of distinct GM composition. HBM feeding promotes the growth of a bifidobacteria-dominated gut microbiome, which exerts a major effect on immune functions (Bäckhed et al., 2015; Stewart et al., 2018; Vandenplas et al., 2020). IF feeding results in the formation of a GM highly diverse but unstable, with overrepresentation of Clostridium difficile (Azad et al., 2013; Jost et al., 2015; Tannock et al., 2013). To mimic the HBM-fed gut microbiome and its beneficial functions, attempts of IF probiotic supplementation have been performed. Lactobacillus rhamnosus GG supplementation in HBM or IF for a month in eight- to ten-month-old infants lowers the proportion of Clostridium histolyticum type bacteria to the total bacterial count (Pärtty et al., 2013). C. histolyticum increases intestinal dysbiosis, which can regulate GI peptide production correlated with satiety, and subsequently increases food intake and the risk of developing obesity (Gomes et al., 2018). Bifidobacterium lactis BB-12 supplementation increases Bifidobacterial abundance, lowers Enterobacteriaceae and Clostridium spp. counts, and exerts no effect on colonization of antibiotic-resistant organisms (Mohan et al., 2006). The presence of Enterobacteriaceae is a hallmark of dysbiosis (Sassone-Corsi et al., 2016), and both Enterobacteriaceae and Clostridium spp. are frequently isolated in cancer patients (Touchefeu et al., 2014).
However, the results of controlling GM by probiotic supplementation remain controversial. Probiotic supplementation in IF failed to exert any considerable effect on growth, stool consistency, incidence of diarrhea, crying, or vomiting (Mugambi et al., 2012). The European Food Safety Authority (EFSA) states that more scientific evidence is necessary to prove the existence of a relationship between probiotic product consumption and their health-related beneficial effects (Rijkers et al., 2011). Thus, more clinical evidence is warranted to elucidate the effects of probiotic supplementation on infant growth and GM composition.
Therefore, our study aimed to improve infant gut health and growth performance by providing probiotic-fortified infant formula (PFIF) and by determining its effects on infant GM. Infant participants (n=118) were allocated into the following three groups: breast milk-fed group (BF), probiotic-fortified formula-fed group (PF), and placebo formula-fed group (FF). Beginning from 30 days postnatal, each designated diet was provided for a period of 4 weeks, and during this period, fecal samples were collected three times a week. Effects of the diet were determined by analyzing fecal consistency, pH, short-chain fatty acid (SCFA) levels, secretory immunoglobulin A (sIgA) levels, and fecal microbiota composition by next-generation sequencing.
Materials and Methods
To conduct the present study, participating mothers and infants were recruited from postnatal care centers (Seoul, Korea). The aims, protocol, and outcomes of the present study were explained to all participants, and they were inquired about their willingness to participate. Exclusion criteria were as follows: severe disorder of the liver, heart, respiratory organs, endocrine glands, or metabolism; and use of antibiotic medication at the time of the clinical trial. A total of 118 participants were recruited for this study. The study was approved by the Korea University Institutional Review Board (IRB, 1040548-KU-IRB-17-251-A-2, and KUIRB-2019-0136-01), which is independent of the Lotte Food Written informed consent was obtained from all participants.
The IF and bacterial strains used in this study were obtained from the Dairy Department of Lotte R&D Center (Seoul, Korea). L. rhamnosus GG, Lactobacillus fermentum LC40, Lactobacillus reuteri DSM 17938, and B. lactis BB12 were selected based on their acid, bile tolerance, and safety assays. Strains were cultured in Man Rogosa Sharpe broth (MRS; Difco, Detroit, MI, USA) for 18 h at 37°C. The strains were subcultured three times prior to the conduction of each experiment. All strains were prepared using 50 % (v/v) glycerol as a cryoprotectant and maintained at –80°C until use. Infants consumed approximately 1.34×107 CFU/mL probiotics in formula without the presence of any other bacterial species.
This randomized, double-blind, placebo-controlled study was performed from January 1, 2018, to June 1, 2020. Randomization was conducted using a computer program, and participants were assigned into the following three groups: BF, PF, and FF. Individual differences in gestational period and infant body weight were minimized among the three groups. Each feeding was practiced for a period of 4 weeks, beginning from 30 days postnatally. Participants received stool containers with a spatula placed inside (SPL Life Science, Pocheon, Korea), together with a sanitary plastic bag, instruction form, and a questionnaire. Participants collected stool samples from the diaper into the collection tube and sent it directly to the microbiology lab (Seoul, Korea) via an express courier. Participants were recommended to collect and send samples early in the morning or late evening to avoid delays.
Both the weight and height of all infants were measured on the following three designated days: 1st, day of birth, 2nd; second, 1 month after the birth; and 3rd, last day of the experiment. The measurements of both weight and height were conducted in triplicate.
Infant fecal samples were collected by their parents immediately after defecation. Concurrently, the defecation frequency was determined following the instructions provided in a paper format. The samples were collected on three different days in one week. Collected samples were then stored at 80°C until analysis in the microbiology lab (Seoul, Korea).
Following the protocol prescribed by the Bristol stool chart, three of our lab members performed fecal consistency analysis. The thawed fecal samples were randomly distributed to all participating members at one time, and the samples were scored.
Fecal pH was measured to determine whether the treatments could change the acidity of the feces in the gut. The weight of each sample was measured to 1 g prior to the conduction of measurements, and the samples were then placed into 15-mL tubes (SPL Life Science) with saline. The pH of the mixed fecal samples was measured using a pH meter (Agilent Technologies, Santa Clara, CA, USA).
Fecal samples were weighed individually in an Eppendorf tube according to the methods reported by Eor et al. (2020); 10 mg fecal samples in Eppendorf tubes were included for analysis individually. The samples were then homogenized after addition of 100 μL crotonic acid, 50 μL HCl, and 200 μL ether. Homogenates were then centrifuged at 1,000×g for 10 min. After centrifugation, the top ether layer was transferred to glass vials. The collected ether was then mixed with 16 μL N-tert-butyldimethylsilyl-N-methylttrifluoroacetamide (MTBSTFA), and the vials were sealed with ParafilmM (Sigma-Aldrich, St. Louis, MO, USA). The samples were heated at 80°C for 20 min in a water bath and then incubated for 48 h. The samples were then placed into the 6890N Network GC system (Agilent Technologies, Wilmington, DE, USA) with an HP-5MS column (0.25 mm×330 mm×30.25 mm) and 5973 Network Mass Selective Detector (Agilent Technologies). Helium (purity 99.9999%) was used as a delivery gas and the carrier gas flow rate was 1.2 mL per min. The head pressure was 97 kPa and split at 20:1. The inlet and temperature of the transfer line were 250°C and 260°C, respectively. The following temperature program was used: 60°C (3 min), 60°C–120°C (5°C/min), and 120°C–300°C (20°C/min). One microliter of the sample was injected (run time: 30 min). SCFA concentrations were then determined by comparing their peak areas with the standards.
S-IgA was quantified using the Secretory IgA ELISA kit (ImmuChrom, Heppenheim, Germany). All procedures were performed according to the manufacturer’s instructions. Stool samples (100 mg) were mixed with 5 mL of wash buffer on a vortex mixer until a homogenous mixture was obtained. One milliliter of the mixture was then transferred into an “Eppendorf” reaction vial and centrifuged for 10 min at 2,000×g. The supernatant was diluted 1:250 with wash buffer (4 μL+996 μL wash buffer). One hundred microliters of the dilution was used per well. The sample preparation method was performed for a total of four times for each sample.
To determine the intestinal microbiota composition, genomic DNA (gDNA) was extracted from the stool samples of each group using the QIAamp DNA stool kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Briefly, 200 mg of fecal sample was mixed with 1.4 mL ASL buffer in a 2-mL tube and vortexed until the sample was thoroughly homogenized. Samples were subsequently heated for 5 min at 95°C, vortexed for 15 s, and centrifuged at 196,768×g for 1 min to pellet the stool particles. After 1.2 mL of the supernatant was transferred into a new 2-mL tube, 1 InhibitEX Tablet was added, and the tube was vortexed immediately until the tablet was completely suspended. The tube was incubated for 1 min at 25°C, centrifuged for 3 min at full speed, and 200 μL of supernatant was transferred into a new 1.5-mL tube. After 15 μL proteinase K and 200 μL Buffer AL were added and the mixture was vortexed, the tube containing the mixture was incubated at 70°C for 10 min. An additional 200 μL of ethanol was added to the lysate and mixed by vortexing. The complete lysate was transferred to a QIAamp spin column with a 2-mL collection tube and centrifuged for 1 min. 500 μL Buffer AW1 was added, and the column was centrifuged at full speed for 1 min. The collection tube containing the filtrate was then discarded. The same process was conducted with 500 μL of AW2. Finally, the QIAamp spin column was transferred into a new 1.5-mL tube, and 200 μL of Buffer AE was directly pipetted onto the QIAamp membrane. All eluted DNA samples were acquired from the collection tube after 1 min of centrifugation.
Small subunit (16S) ribosomal DNA amplicon sequencing was performed to analyze the microbial community structure. The hyper variable V3-V4 regions of 16S rDNA were amplified using the following universal primer set (forward 5′-CCT-ACG-GGN-GGC-WGC-AG-3′, reverse 5′-GAC-TAC-HVG-GGT-ATC-TAA-TCC-3′). The resulting DNA amplicon of ~460 bp was purified using the Agencourt AMPure XP kit (Beckman Coulter, Brea, CA, USA). The second PCR was performed to attach Illumina universal p5/p7 overhang sequences and sample-specific barcodes. A sequencing library of ~550 bp was purified using the Agencourt AMPure XP kit (Beckman Coulter). Sequencing was conducted using the Illumina MiSeq platform (Illumina, San Diego, CA, USA) using the MiSeq Reagent kit V3 (2×300 PE; Illumina, San Diego, CA, USA). Control experiments for DNA extraction, library preparation, and sequencing were performed as follows. The mock DNA library was prepared using the ZymoBiomics microbial community DNA standard (Zymo Research, Irvine, CA, USA) using the DNA amplicon library preparation procedure described above. Negative control was established by following the same DNA extraction procedure without feces. The final elute of the DNA extraction negative control was used as the template for PCR negative control, following the same PCR conditions described above. We confirmed that the samples were not contaminated during DNA extraction or library preparation.
Sequencing reads were processed using the DADA2 pipeline (Ver 1.12.1). Reads were trimmed and filtered, paired reads were merged, and chimeric sequences were removed. The SILVA ribosomal RNA reference database (Release 128) was used for taxonomic assignment of the amplicon sequence variants. Data on low-quality samples with <10,000 observed operational taxonomic units (OTUs) were removed from the sample set. Among the 1,984 total OTUs, only 762 OTUs with a total count of more than 100 were considered. The microbiome R/Bioconductor package was used to calculate the alpha diversity. The community structure was illustrated using an in-house R script based on the phyloseq R/Bionconductor package.
IBM SPSS Statistics software (version 25.0; IBM, Armonk, NY, USA) was used to analyze the data. One-way analysis of variance was used to compare the sample means. Multiple comparisons of means were performed using the Duncan’s multiple range test and Tukey’s post-hoc test. p<0.05 was considered statistically significant.
Results
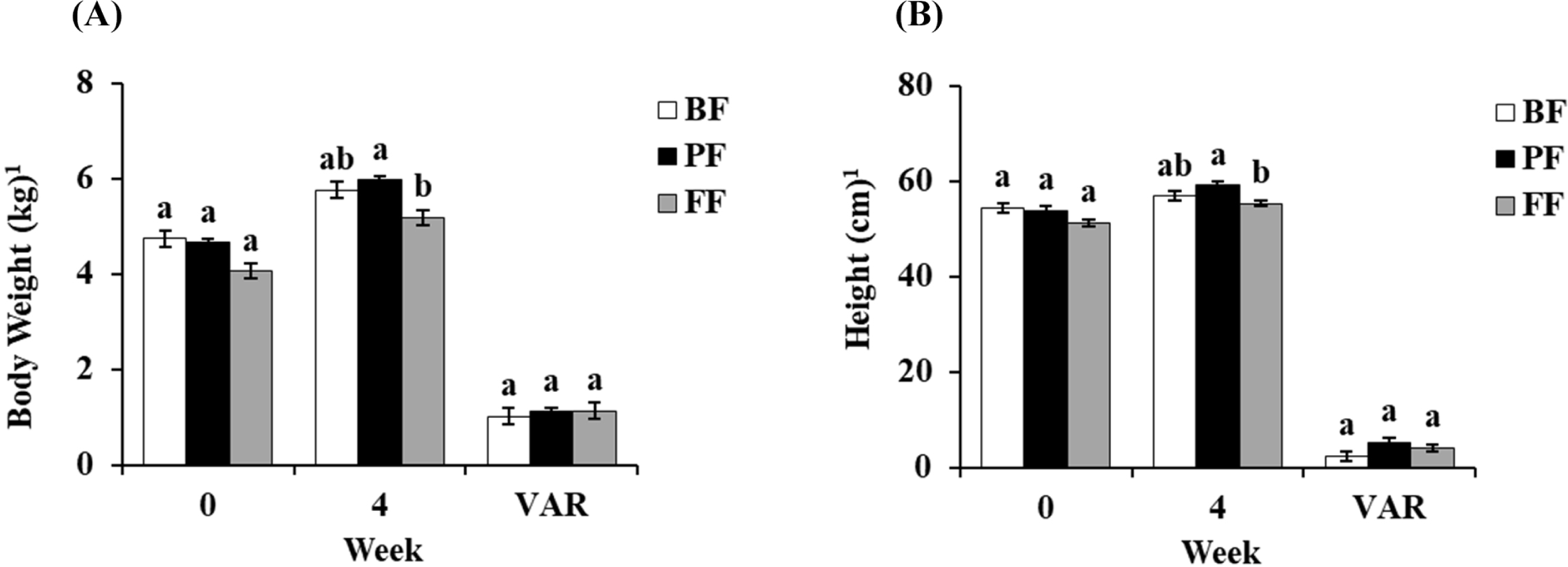
A slight change in body weight was observed after each treatment. The initial body weights of BF, PF, and FF at 0 weeks were 4.74, 4.65, and 4.06 kg. After 4 weeks of treatment, the body weights of three groups were increased to 5.75, 5.97, and 5.18 kg. Although the difference between the treatment periods were 1.01, 1.11, and 1.13 kg. Significantly increased body weight was observed in the 4th week of PF treatment compared to FF (p<0.05; Fig. 1A). Moreover, similar to the observations with body weight increase, the heights of PF groups was also significantly higher than FF treated groups (p<0.05), which indicates the increased growth performance of PF group (Fig. 1B). The heights in groups BF, PF, and FF at 0 weeks were 54.4, 53.9, and 51.2 cm. However, in the 4th week, the heights were 56.8, 59.1, and 55.3 cm, with a variance (VAR) within 4 weeks showing 2.4, 5.3, and 4.1 cm, respectively.
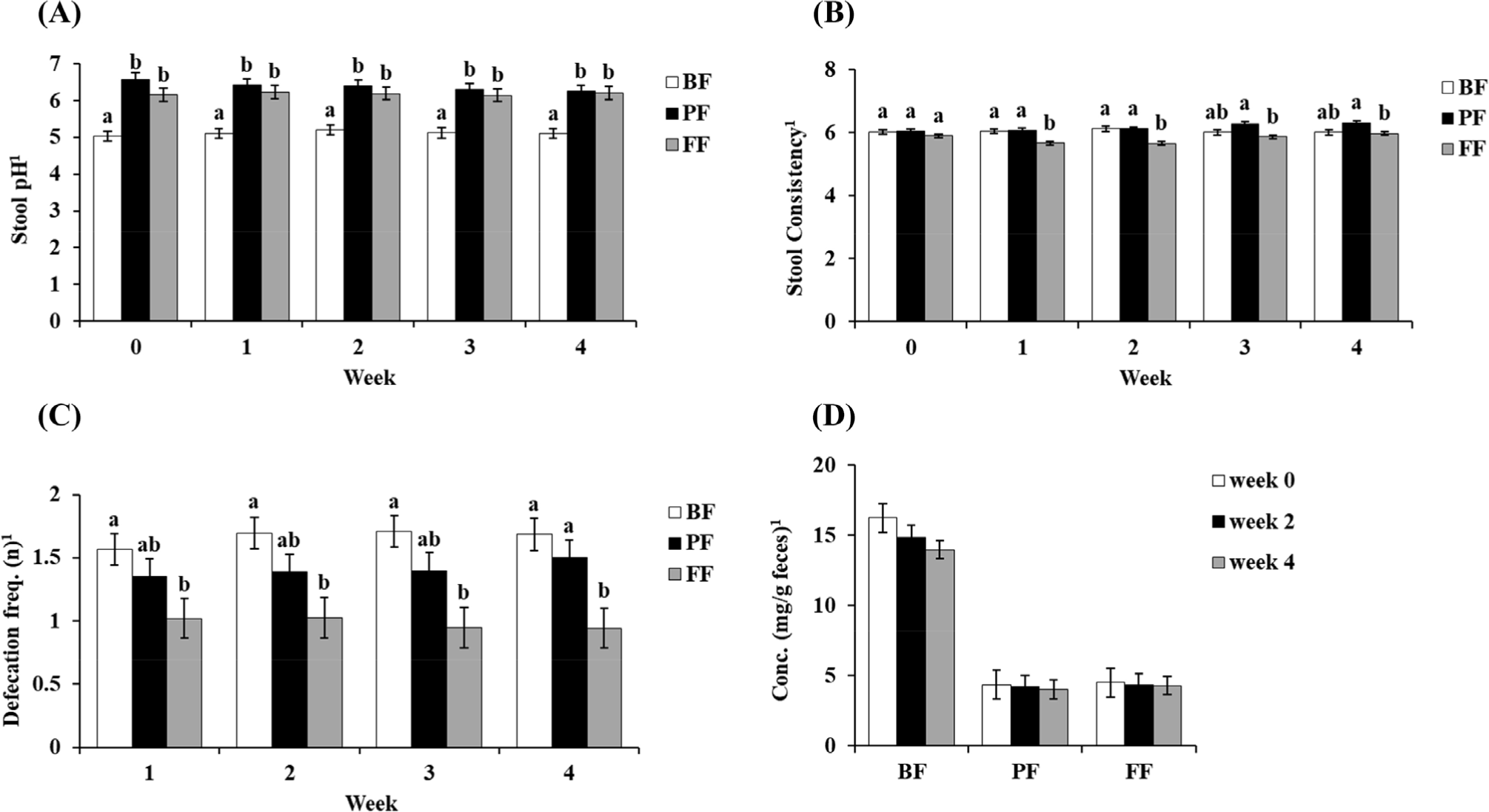
The four types of assessment of stool conditions are illustrated in Fig. 2. First, the results of stool pH analysis showed significantly lower levels in the BF groups compared to other groups (p<0.05; Fig. 2A). Second, stool consistency was analyzed by scoring the condition of the stool samples using the Bristol chart method (Fig. 2B). The results suggested that stool consistency did not considerably changed in the BF, PF, and FF groups throughout the study period. However, the average consistency of FF showed a significantly decreased score rate compared to BF and PF groups since 1st week to the study period (Fig. 2B). The frequency of defecation showed the same trend from the 1st week to the 3rd week. However, the 4th week results indicated that FF showed a significant decrease defecation frequency score (p<0.05) compared to both the BF and PF groups (Fig. 2C). Finally, the stool IgA levels in each treatment throughout the study period did not show any significant difference (Fig. 2D).
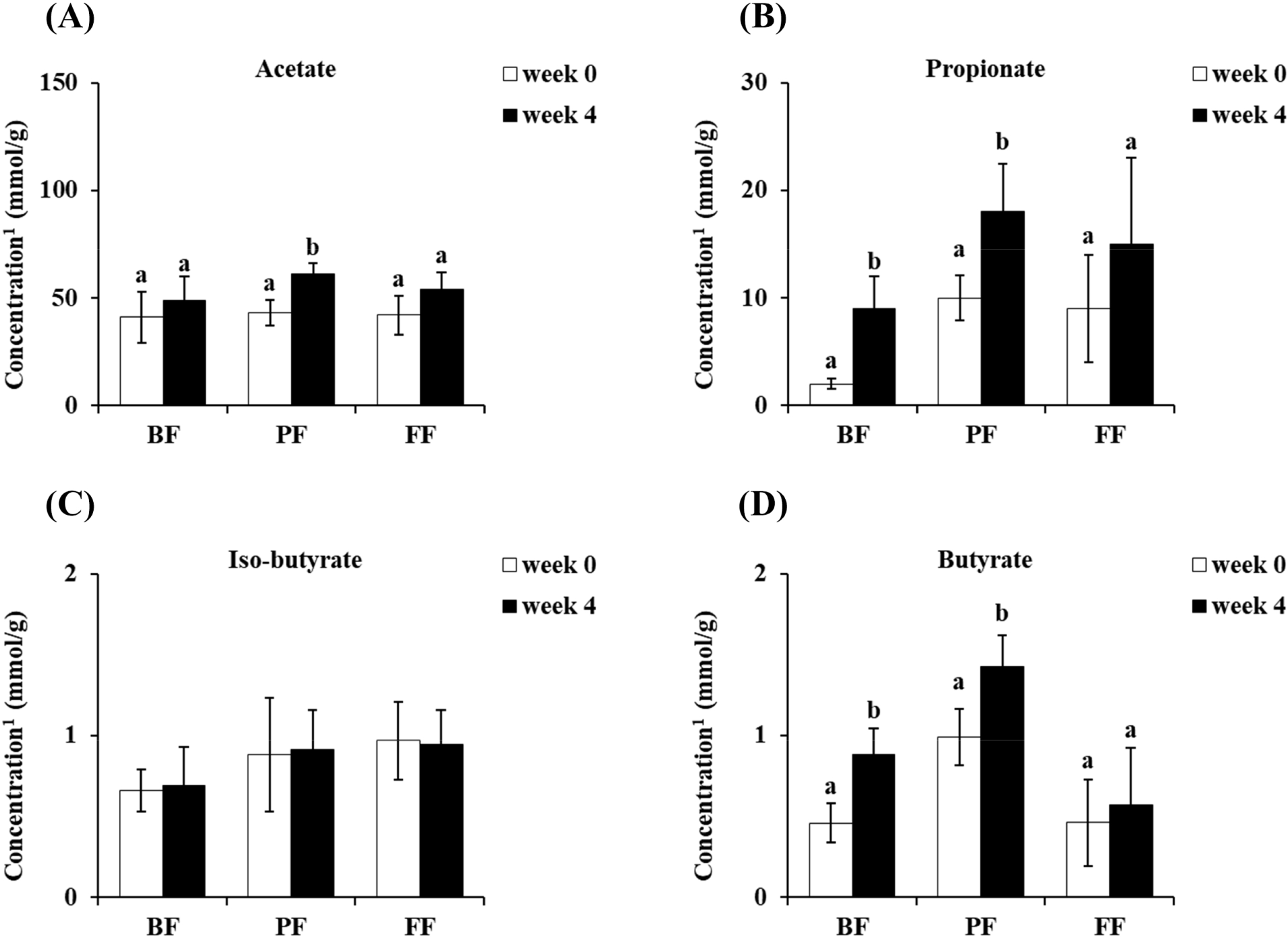
Stool SCFAs were analyzed before and after the conduction of each treatment. The concentration of acetate showed a significant increase only in the PF group (p<0.05; Fig. 3A). Additionally, both the BF and PF groups showed significant increases (p<0.05) in terms of both propionate and butyrate concentration after 4 weeks of treatment (Figs. 3B and D). However, the iso-butyrate concentration showed no considerable difference after 4 weeks of treatment (Fig. 3C).
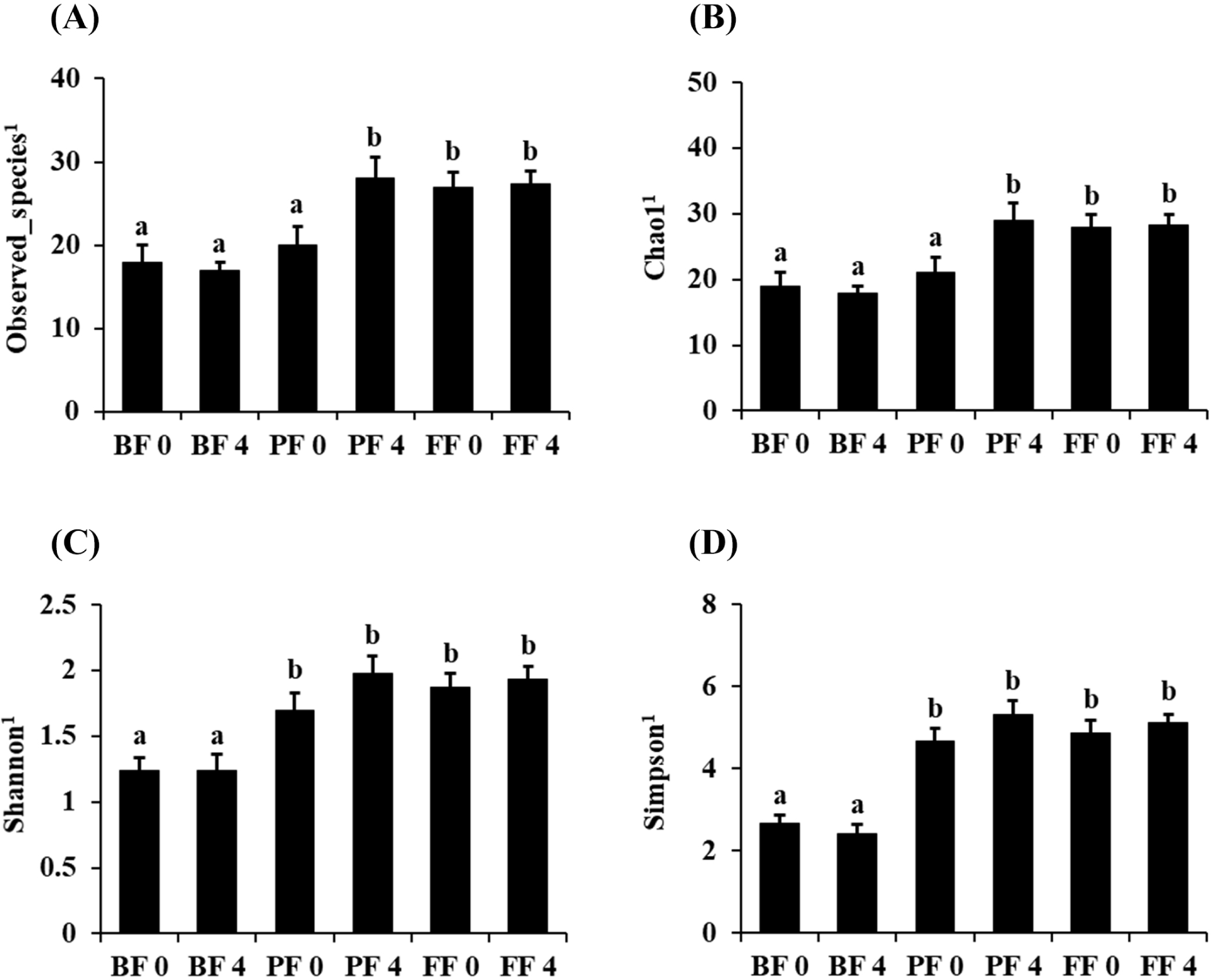
Fecal microbiota alpha diversity was calculated using the observed OTUs, Chao1, Shannon, and Simpson (Fig. 4). Overall, there were remarkable differences between the BF and PF groups after 4 weeks of each treatment in terms of the OTUs (Fig. 4A), Chao1 (Fig. 4B), Shannon (Fig. 4C), and Simpson (Fig. 4D) indices. The alpha diversity changes in each treatment group showed no significant differences between 0 and 4 weeks after treatment, except in the PF group (Figs. 4A and B). Furthermore, the Shannon and Simpson indices indicated significant increased diversity between BF and PF after 4 weeks of treatment (p<0.05; Figs. 4C and D). The results indicate that PF and FF treatment are capable of increasing the diversity of observed species in infant gut.
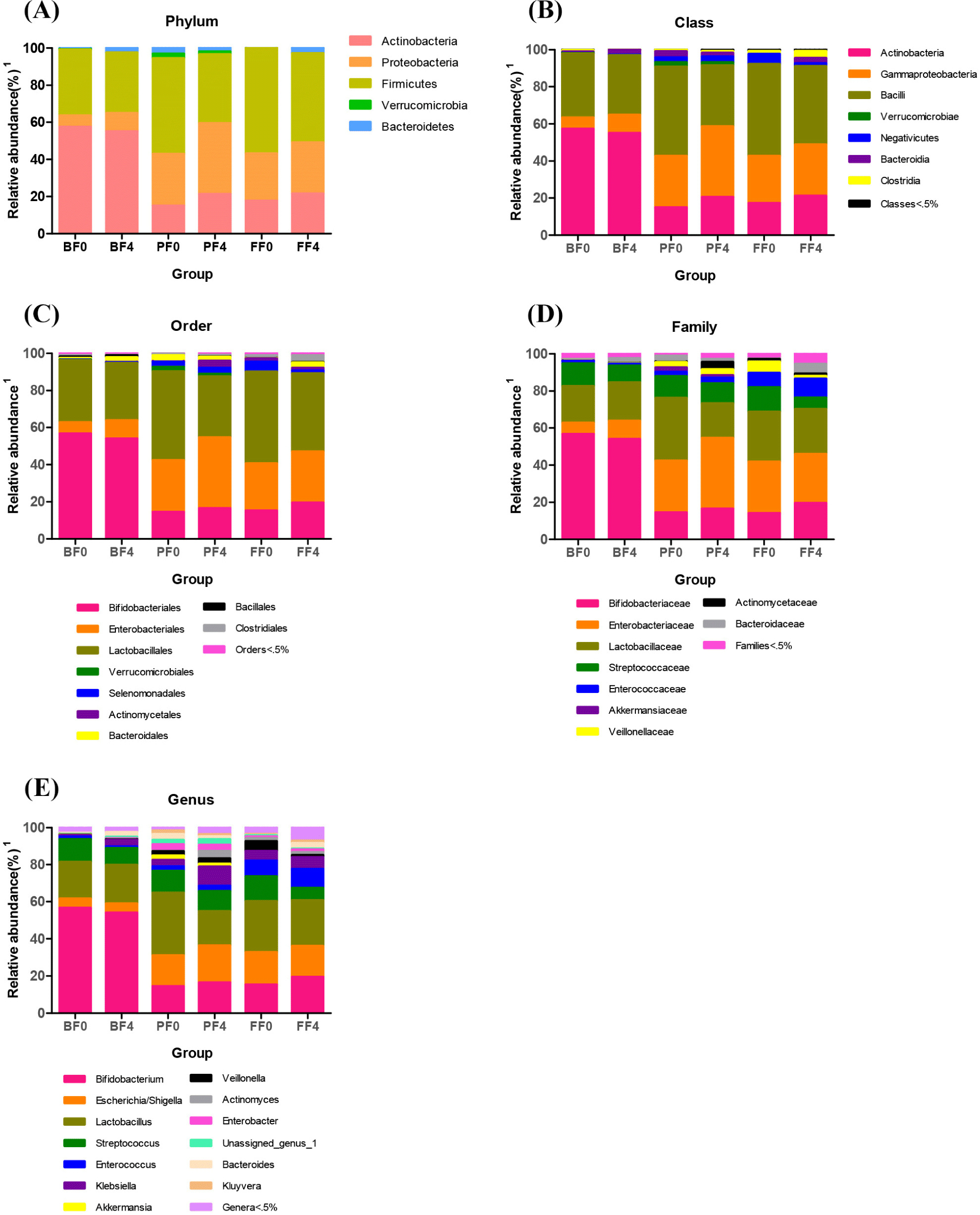
At various taxonomic levels, gut microbiome composition was compared between the 0- and 24-week fecal samples in each treatment group. Overall, there were no substantial differences in the microbiome composition changes at various levels in the BF group (Fig. 5). At the phylum level, Firmicutes showed significantly less (p<0.05) abundance in the gut microbiome in the 4-week samples than that in the 0-day samples in the PF and FF groups. However, the abundance of Actinobacteria and Proteobacteria increased after 4 weeks of treatment (Fig. 5A). At the class level, Bacilli showed significantly less (p<0.05) abundance in the GM at 4 weeks in the PF and FF groups. However, the relative abundances of Gamma proteobacteria and Actinobacteria increased after 4 weeks (Fig. 5B). At the order level, the abundance of Lactobacillales was reduced in the 4-week samples in the PF and FF groups. However, Enterobacteriales expanded after 4 weeks of treatment (Fig. 5C). At the family level, Lactobacillaceae abundance was reduced after 4 weeks of treatment in the PF and FF groups. However, the relative abundances of Enterobacteriaceae and Bifidobacteriaceae increased significantly (p<0.05) at 4 weeks (Fig. 5D). At the genus level, Lactobacillus abundance decreased after 4 weeks of treatment in the PF and FF groups. Notably, Streptococcus abundance reduced in the 4th week samples in the FF group. However, Escherichia, Shigella, and Bifidobacterium abundances increased significantly (p<0.05) after 4 weeks of treatment (Fig. 5E).
Discussion
A considerable height change in PF and FF was observed in the 4th week of treatment. Importantly, the variance of height showed an increase in both PF and FF compared to BF. The PFIF showed remarkable differences in height and weight compared to the BF and FF groups. This result was consistent with the results reported by a previous study conducted by Vendt et al. (2006).
The difference in stool pH shows an alteration in the GI environment, and a lower pH of stool may suggest a more enhanced state of the GI tract occurring due to pathogens (Henrick et al., 2018). Therefore, the BF group reveals the most protective effect conferred against viral infection and demonstrates the effect exerted on the immune system of newborn babies against infection among the group (Moore et al., 2021; Toscano et al., 2017). Stool consistency, which indicates the state of stools expelled by the body, showed a significant difference (p<0.05) occurring since the 1st week, and PF restored the decrease and maintained almost the same results as BF. The stool defecation frequency indicates the motility and healthiness of the gut. The results showed a significant difference (p<0.05) occurring since the 1st week to the 4th week without critical changes. These results were similar to those reported by previous studies conducted using probiotics and prebiotics, respectively (Vandenplas et al., 2015; Vendt et al., 2006). Stool IgA concentration did not change in each week. These results were similar to those reported by a previous study (Bakker‐Zierikzee et al., 2006).
Based on the GM modulation, changes in each level indicated that dietary differences might induce microbiota changes. At the phylum level, we identified the relative abundances of Actinobacteria, Proteobacteria, Firmicutes, Verrucomicrobia, and Bacteroidetes. In our study, the lower abundance of Firmicutes, Verrucomicrobia, Bacteroidetes, and higher abundance of Proteobacteria and Actinobacteria were associated with the treatment effects of probiotics and the formulation. In comparison, data obtained from human and animal studies showed that the abundance of Firmicutes and Actinobacteria increased in obesity models and tended to increase with weight gain. Moreover, the proportions of Bacteroidetes and Verrucomicrobia are decreased in obese subjects and the rate of obesity continues to increase (Chakraborti, 2015; Zhang et al., 2021). However, the results of the present study were different from those of previous studies conducted on body weight and height. Furthermore, the chief butyrate-producing bacteria in the human gut belong to the phylum Firmicutes (Louis and Flint, 2009; Louis and Flint, 2017). Butyrate is a SCFA derived from the microbial fermentation of dietary fibers in the colon and it is recognized for its potential to act on secondary chemoprevention by slowing growth and activating apoptosis in colon cancer cells (Berni Canani et al., 2012). Actinobacteria, Bacteroidetes, and Proteobacteria are potential butyrate producers (Vital et al., 2014). Additionally, apart from butyrate, the production of other SCFAs is mediated by bacterial members belonging to Actinobacteria that produce acetate and lactate during carbohydrate fermentation (Rivière et al., 2016). Also, the mucin-degrading bacteria phylum Verrucomicrobia produces both propionate and acetate (Derrien et al., 2004). In this study, we revealed that the relative abundances of Actinobacteria and Proteobacteria were significantly increased (p<0.05) in the probiotics and formula-fed groups. Our results are consistent with those reported by previous studies. In stool consistency and defecation frequency analyses, several studies have reported that Bacteroidetes to Firmicutes ratio is higher in the subjects with a small amount of defecation than in those with a considerable amount of defecation, and at the genus level, Bifidobacterium is less abundant in the subjects with a small amount of defecation (Kwon et al., 2019). Moreover, remarkable increase in the relative abundance of Firmicutes in the gut microbiome of group presenting with loose stool in comparison with the group presenting with normal stool consistency (Bragg et al., 2020). In this study, the results of stool consistency were similar to those reported by previous studies. The stool consistency and stool defecation frequency rates were significantly higher (p<0.05) in BF fed group compared to FF group, which can indicated the gut health of the infants. This result is similar with the study reported by Vandeputte et al. (2016). and the colleges which have shown that increased gut health was correlated with the increased diversity of GM (Vandeputte et al., 2016). Compared to FF group, BF and PF fed groups have shown the increased diversity of GM in the genus level was observed in our study. Moreover, at the genus level, we discovered that the relative abundances of Bifidobacterium, Escherichia/Shigella, Streptococcus, Klebsiella, and Bacteroides were considerably different in the fecal microbiota of the probiotic and formula-fed groups. Notably, previous studies indicated that a decreased abundance of Bacteroides was associated with the development of several diseases, such as obesity and diabetes (Bervoets et al., 2013; Zhang et al., 2013). In our study, the results are similar to those reported by previous studies in terms of the probiotic formula-fed group with body weight gain. Moreover, the relative abundance of the genus Bacteroides demonstrates a high capacity for the production of SCFAs such as butyrate (Ohira et al., 2017).
Conclusion
The present study showed overall fecal microbiota modulation induced by dietary changes in infants. Although the results from BF intestinal health promoting effect showed the better than FF groups, probiotic strain fortified FF showed improved effect on gut health. Based on the effects, we analyzed GM from the fecal samples. The GM composition and the abundance of health-promoting bacteria increased in the probiotic formula group compared to the FF. Supplementation with probiotics can help optimize the infant gut environment, composition, and metabolic activity of the microbiota, mimicking those of breast milk. Despite this limitation exist, we tried to find overall meaning from changed genera that have previously been reported. Additionally, the findings from this study would provide strategy or reference who wants to investigate on manufacturing fortified IF for infants with novel GM modulation.