










Introduction
Proteolysis is an enzymatic reaction involving multiple systems in which proteins are broken down into peptides or amino acids. Proteolysis plays an important role in controlling cell metabolic processes, such as cell division, apoptosis, transcription, signal transduction, protein degradation, and the regulation of multiple metabolic pathways (Ciechanover, 2005). Proteolysis also occurs in meat and is involved in vital activities of a series of enzymatic reactions (Barido and Lee, 2021; Lana and Zolla, 2016). Proteolysis mainly involves different endogenous proteolytic enzyme systems, such as calpains, cathepsins, proteasome, and caspases (Koohmaraie, 1996; Lana and Zolla, 2016; Sentandreu et al., 2002). The mechanism of proteolysis in beef and pork during storage has been widely elucidated (Lametsch and Bendixen, 2001; Lametsch et al., 2003; Lana and Zolla, 2016). However, this has not been fully studied in poultry meat, especially duck meat, due to the relatively little attention paid to poultry tenderization compared to that of beef or pork.
Recently, new insights into tenderness and meat color changes through proteomics and the importance of proteomics research in postmortem metabolic activities have been proposed (Purslow et al., 2021). Previous studies have demonstrated the proteome change and its relationship with beef or pork quality change postmortem (Gagaoua et al., 2020; Gagaoua et al., 2021; Jia et al., 2006; Lametsch and Bendixen, 2001; Lametsch et al., 2003). Duck meat, along with chicken and turkey meat, is classified as poultry meat, but its physicochemical properties and muscle fiber composition are different from those of other poultry meat (Kim et al., 2008). Thus, the differences in the trend of proteolysis between duck meat and chicken meat are expected due to the differences in the characteristics of muscle fiber and physicochemical properties between duck and chicken meat. However, until now, proteome change from a proteolysis perspective and its relation to meat quality characteristics in duck meat has not been fully investigated.
Therefore, the purpose of this study was to investigate protein degradation by proteolysis during cold storage in two types of duck skeletal muscle (M. pectoralis major; PM and M. iliotibialis, IL) that have different physiological functions and different muscle fiber types. In addition, the relationship between proteolysis and meat quality characteristics was investigated to better understand the postmortem physicochemical changes in duck meat.
Materials and Methods
Fifteen ducks were slaughtered at a commercial slaughterhouse according to the standard commercial procedures of the Korean livestock production system. Duck PM (n=30) and IL (n=30) muscles were taken from both right and left sides of carcasses (6-week-old Cherry Valley ducks, 2.4±0.3 kg) at 6 h postmortem. Muscles were weighed individually, vacuum-packed (1.0 bar; MVAC 300, Maxima, Netherlands), and stored in a cold room at 4°C. Ten PM and IL muscles each were randomly selected at different storage times (1, 5, and 10 days), respectively. The meat quality characteristics, proteolytic enzyme activity, and proteolysis-induced peptides of each individual muscle were analyzed at different storage times. For analysis of proximate composition (moisture, crude protein, crude fat, and crude ash) and muscle fiber characteristics, samples were collected from both muscles on day 1.
Moisture, crude protein, and crude ash contents were analyzed using the AOAC (2000) method. Crude fat content was determined using the Folch et al. (1957) method with some modifications. Briefly, three grams of the sample were homogenized with 20 mL of Folch solution (chloroform: methanol, 2:1, v/v) at 8,000 rpm for 20 s using a homogenizer (T18, IKA Works GmbH & Co., Staufen, Germany). After filtrating the homogenate using filter paper (Whatman No. 1; Merck KGaA, Darmstadt, Germany), the filtrates were separated into two layers using 0.88% NaCl. The lower layer was collected, and the solvent was removed using nitrogen gas. Crude fat content was presented as a percentage of the weight of the sample. Proximate composition evaluation was conducted in triplicate for each sample.
Muscle pH was measured with homogenates of 3 g samples and 27 mL deionized water using a pH meter (MP230, Mettler-Toledo, Greifensee, Switzerland) calibrated with standard buffers (pH 4.01, 7.00, and 9.21). Meat color was measured using a colorimeter (CR-400, Minolta, Tokyo, Japan) calibrated with a white plate (Y=93.5, x=0.3132, y=0.3198). A Commission Internationale de l’Eclairage (CIE, 1978) system was used to determine the color values (CIE L*, lightness; CIE a*, redness; CIE b*; yellowness). Purge loss and cooking loss were measured to determine water-holding capacity. To analyze the purge loss, each muscle was removed from the package and weighed at different storage times. The weight difference before and after storage was recorded as purge loss (%). To determine the cooking loss of the sample, muscles were cooked in a water bath at 75°C until their internal temperature reached 70°C. The weight difference before and after cooking was recorded as the cooking loss (%). The cooked samples were used to measure shear force. Three cores were removed from each sample by cutting parallel to the muscle fiber direction. Each core was cut vertically using a texture analyzer (TA1, Ametek, Largo, FL, USA) equipped with a Warner-Bratzler shear blade. The Warner-Bratzler shear force (WBSF; kg/cm2) value was recorded for each sample with the average of the three cores.
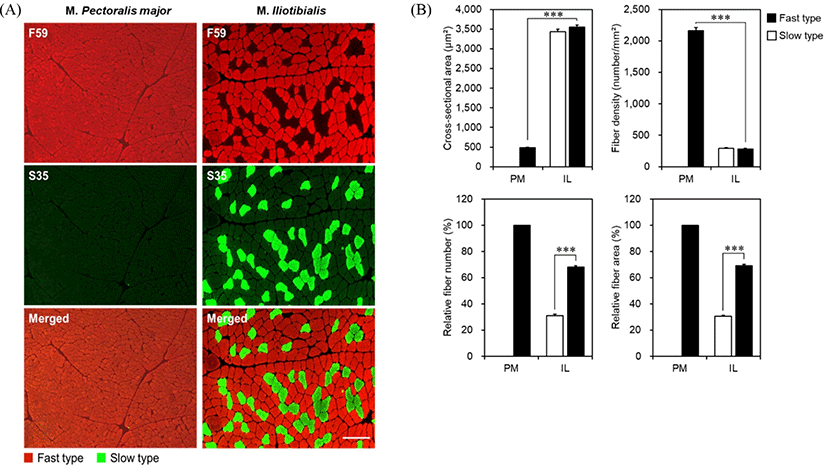
To analyze muscle fiber characteristics, the muscle fibers of duck PM and IL muscles were cut into small pieces (0.5×0.5×0.5 cm) and immediately frozen using 2-methylbutane chilled with liquid nitrogen. Muscle fiber staining was performed using the Song et al. (2020) method with some modifications. Briefly, transverse sections (10°Cμm in thickness) were obtained from the frozen muscles using a cryostat microtome (CM 1860, Leica Biosystems, Nussloch, Germany) at –21°C. The primary antibodies (F59 and S35; DSHB Iowa, IA, USA) specific to each myosin heavy chain (MHC; slow and fast MHCs, respectively) were applied to the section. To visualize the muscle fibers that reacted with primary antibodies, fluorescent dye-conjugated anti-mouse IgG (Alexa Fluor 594, Thermo Fisher Scientific, Waltham, MA, USA) and IgG2a (Alexa Fluor 488, Thermo Fisher Scientific) were used. Stained sections were captured using a fluorescence microscope (EVOS M5000, Thermo Fisher Scientific), and approximately 800 muscle fibers were counted. The cross-sectional area (CSA, μm2) of each muscle fiber, relative area (%), relative number (%), and fiber density (number/mm2) were analyzed using an Image Pro Plus Program (Media Cybernetics, Rockville, MD, USA).
For peptide extraction, 5 g of the sample was homogenized with 30 mL of 0.01 N HCl for 3 min in a stomacher (BagMixer®400, Interscience, Saint Nom, France). The homogenate was centrifuged at 10,000×g for 30 min at 4°C. The supernatant was filtered through glass wool, and filtrate was mixed with three volumes of ethanol. After being stored at 4°C for 24 h, the mixture was centrifuged at 10,000×g for 30 min at 4°C, and the supernatant was lyophilized using a vacuum evaporator (SPD1010, Thermo Fisher Scientific, MA, USA). The sample was dissolved with 5 mL of 0.01 N HCl, neutralized to pH 7.0 using NaOH, and filtered through a 0.45-μm nylon membrane filter (Millipore, Bedford, MA, USA). The filtrate was centrifuged in centrifugal filter-containing tubes (Amicon® Ultra-15 Centrifugal Filter Unit, Millipore, MA, USA) at 10,000×g for 1 h.
The identification and quantification of peptides in duck muscles was conducted using the Kim et al. (2021) method with some modifications. Briefly, peptides were analyzed using an LC-MS/MS equipped with an LC device (Easy-n-LC, Thermo Fisher, San Jose, CA, USA) coupled with a C18 nano bore column (150 mm×0.1 mm, pore size of 3 μm, Agilent Technologies, Santa Clara, CA, USA) and an LTQ-Orbitrap XL mass spectrometer (Thermo Fisher, San Jose, CA, USA). The MS/MS spectra were identified and quantified using the PEAKS Studio 10.0 (Bioinformatics Solutions, Waterloo, ON, Canada), and the database was derived from UniProt [release March 2020 from http://www.uniprot.org, taxonomy Anas 8835 (45,681 sequences)].
Three different extraction buffers were prepared to extract proteolytic enzymes from the samples: cathepsins B and L (150 mM NaCl, 25 mM Tris, 50 mM EDTA, 1.0 mM DTT and 1.0% Triton-100, pH=7.6), calpains and proteasome 20S (100 mM Tris, 10 mM DTT, and 10 mM EDTA, pH to 8.3), and caspase-3 (100 mM HEPES, 0.5 mM EDTA, 5 mM DTT, 20% glycerol, and 0.2% SDS, pH 7.5). The extraction of enzymes was done using a modified version of the method by He et al. (2019). A total of 200 mg of the sample was homogenized with 1.5 mL of extraction buffer using a multipurpose mill (MM400, Retsch GmbH, Dusseldorf, Germany) and centrifuged at 12,000×g for 30 min at 4°C. The supernatant was collected, and its concentration was measured using the method by Bradford (1976). Protein concentration was adjusted to 2.0 mg/mL. The activities of cathepsins B and L were measured using a reaction with the buffer containing the substrates (Z-RR-AMC; EMD, Merck, Darmstadt, Germany, and Z-FR-AMC; SC-3136, Santa Cruz Biotechnology, CA, USA, respectively). The absorbance value was recorded at a wavelength of 380 nm/460 nm (excitation/emission) using a microplate reader (SpectraMax iD3, Molecular Devices, San Jose, CA, USA). Activities of other enzymes were analyzed using the Calpain Activity Fluorometric Assay Kit (MAK228, Sigma-Aldrich, St. Louis, MO, USA), 20S Proteasome Activity Assay Kit (APT280, Merck, Darmstadt, Gemany), and the EnzChek® Caspase-3 Assay Kit #1 (E-13183, Molecular Probes, Eugene, OR, USA) according to the manufacturers’ instructions for calpains, proteasome 20S, and caspase-3, respectively. After reactions with substrates, the absorbance values were collected at a wavelength of 380 nm/460 nm (excitation/emission) for calpains and proteasome 20S and 342 nm/441 nm (excitation/emission) for caspase-3. The enzymes’ activities were presented as the absorbance value relative to the control value.
All data were obtained from triplicate experiments per each sample and were expressed as the mean and standard error of 10 muscles in each group (day 1, day 5, and day 10 of cold storage) of each muscle type (PM and IL). Data analysis was performed using SAS 9.4 software (SAS Institute, Cary, NC, USA). The effect of muscle type on proximate composition and muscle fiber characteristic was examined using the t-test, and the effect of storage time and muscle type on meat quality characteristics and protease activity was examined using two-way ANOVA tests. Ducan’s multiple range tests were used to determine statistical significance at p<0.05, p<0.01, and p<0.001. Principal components analysis (PCA) was conducted to evaluate the relationship between proteins, proteases, and meat quality traits. PCA was performed based on the correlation matrix (PRINCOMP procedure).
Results
The results of proximate composition and meat quality characteristics of duck PM and IL are shown in Table 1. There were no significant differences between the PM and IL muscles in the moisture, crude protein, and crude ash content (p>0.05), whereas the crude fat had a greater proportion in IL than in PM muscle (p<0.05). Duck IL had a higher pH and lightness (CIE L*) but lower cooking loss than PM (p<0.05). In addition, PM had a redder color (higher CIE a*) on day 1 and day 5 than that of IL (p<0.05), although they didn’t show difference on day 10 (p>0.05). During the 10-day cold storage, the pH value of PM declined significantly (p<0.05), while the meat quality traits of IL were not affected by storage (p>0.05). In PM, the redness increased from day 1 to day 5, while cooking loss was less on day 10 than on day 5 (p<0.05). Regardless of muscle type, pH, yellowness (CIE b*), and purge loss were affected by 10 days of storage (p<0.05), but no combination effect with the muscle types and storage time was found (p>0.05).
Muscle fibers are classified into two types (slow and fast) based on the distribution of MHCs, as shown in Fig. 1A. Duck PM muscle was composed only of fast type of muscle fiber, whereas IL muscle was composed of both slow and fast types. PM had a smaller cross-sectional area of fast-type fibers than IL muscle fibers (p<0.001; Fig. 1B). PM had a higher fast-type fiber density than IL muscle (p<0.001). There was no significant difference between fast- and slow-type muscle fibers within the IL muscle in terms of cross-sectional area and fiber density (p>0.05). However, muscle fiber compositions (relative fiber number and area) were significantly higher in the fast type than in the slow type within the IL muscle (p<0.001).
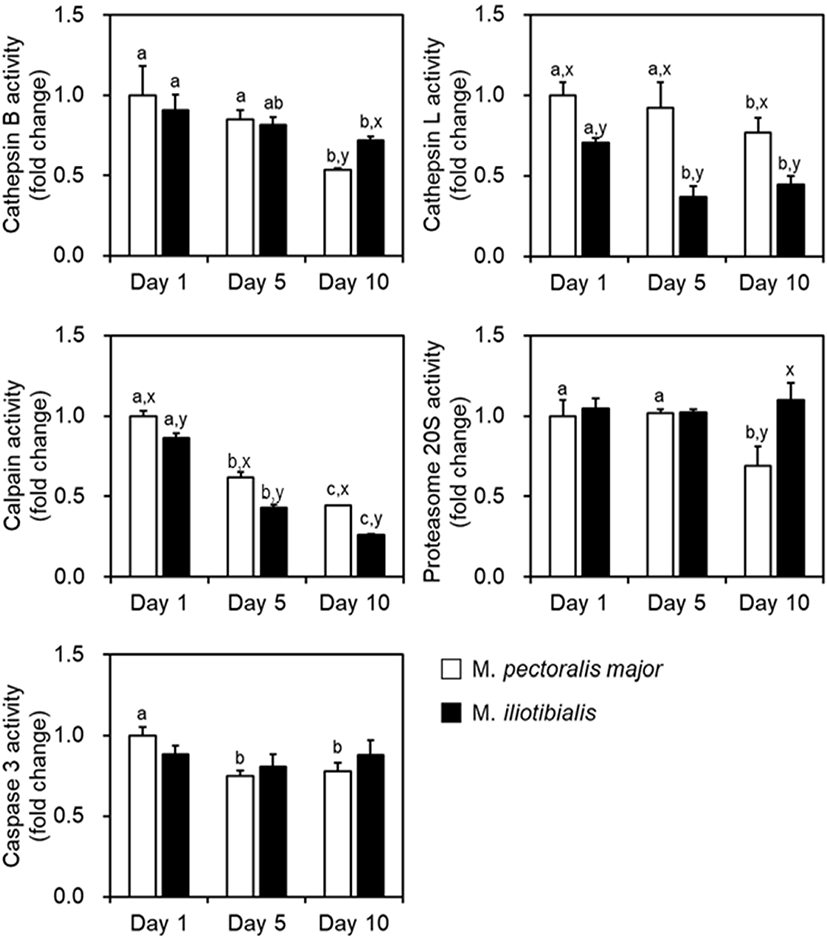
The change in protease activities (calpains, cathepsins L and B, proteasome 20S, and caspase-3) was evaluated during 10 days of cold storage, as presented in Fig. 2. In both muscles, the activity of cathepsins B and L, and calpains decreased after 10 days of cold storage (p<0.05), whereas the activity of proteasome 20S and caspase-3 was decreased only in PM muscle after 5 or 10 days of storage (p<0.05). Duck PM muscle showed higher activity in cathepsin L and calpains than in IL muscle during storage (p<0.05), whereas the activity of cathepsin B and proteasome 20S was higher in PM than in IL only at 10 days of storage (p<0.05). However, the activity of caspase-3 did not show any difference between the two muscles during storage (p>0.05).
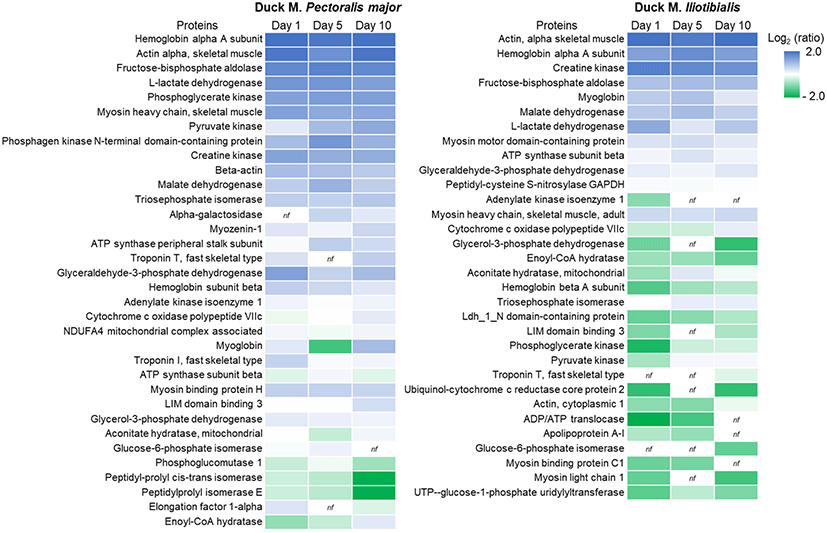
A total of 5,155 peptides were detected and derived from 34 proteins in duck PM, while 4,222 peptides derived from 32 proteins were detected in duck IL (Table 2). Peptides derived from actin, hemoglobin, fructose-bisphosphate aldolase, L-lactate dehydrogenase, MHC, pyruvate kinase, phosphagen kinase N-terminal domain-containing protein, grlyceraldehyde-3-phosphate dehydrogenase, and myoglobin accounted for a large fraction of the total peptides produced by proteolysis in both PM and IL muscle. The heat map (Fig. 3) showed quantitative changes in peptides derived from duck PM and IL muscles during 10 days of storage. As observed in spectral counts (Table 2), hemoglobin, actin, fructose-bisphosphate aldolase, L-lactate dehydrogenase, and creatine kinase showed strong intensities from day 1 regardless of muscle types. The strength of peptides derived from pyruvate kinase, triosephosphate isomerase, and cytochrome c oxidase polypeptide VIIc exhibited a similar tendency in both muscles and increased during 10 days of storage. However, degradation of glyceraldehyde-3-phosphate dehydrogenase, hemoglobin subunit beta, myoglobin, and glycerol-3-phosphate dehydrogenase showed different trends between PM and IL muscles: peptide intensities decreased in PM but increased in IL after 5 or 10 days of storage. For PM, alpha-galactosidase-derived peptides were not initially observed, but peptides were produced after 5 days of storage from alpha-galactosidase. In contrast, no peptides produced from glucose-6-phosphate isomerase after 10 days of storage. In IL muscle, peptides derived from adenylate kinase isoenzyme 1, adenosine diphosphate (ADP)/adenosine triphosphate (ATP) translocase, apolipoprotein A-1, and myosin binding protein C1 were not detected after 10 days of storage, whereas peptides derived from troponin T and glucose-6-phosphate isomerase were observed on day 10.
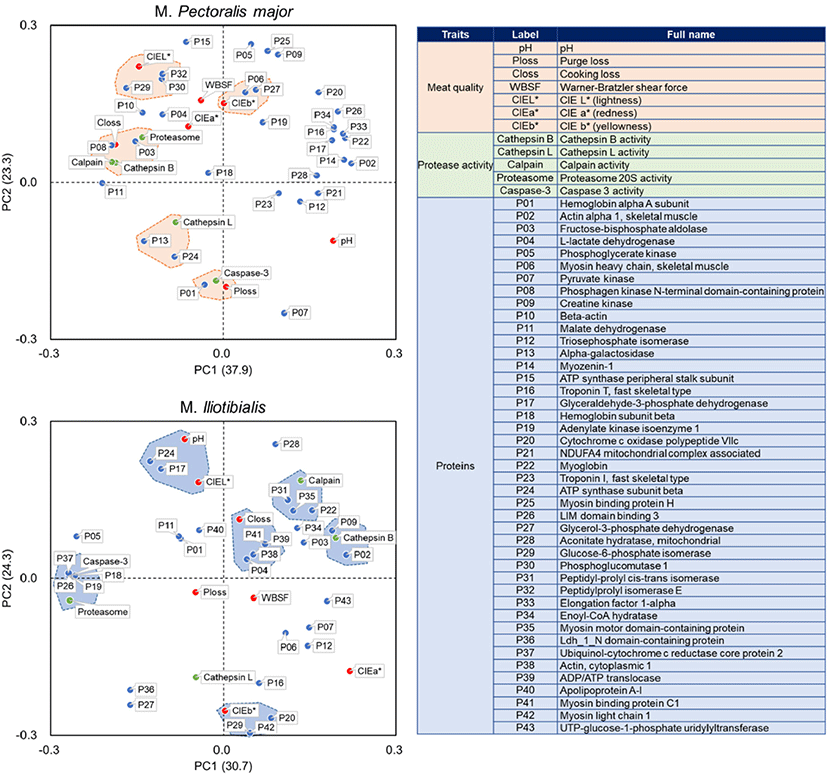
PCA results are shown in Fig. 4 to evaluate the relationships between proteins, proteases, and meat quality characteristics in duck PM and IL muscles. For PM muscle, glucose-6-phosphate isomerase, phosphoglucomutase 1, and peptidylprolyl isomerase E were closely related to lightness (CIE L*), while MHC and glycerol-3-phosphate dehydrogenase were associated with yellowness (CIE b*). Hemoglobin alpha a subunit was linked to purge loss and caspase-3, whereas fructose-bisphosphate aldolase and phosphagen kinase N-terminal domain-containing protein were linked to calpain, cathepsin B, proteasome 20S, and cooking loss. Alpha-glucosidase and ATP synthase subunit beta showed a relationship with cathepsin L. However, these proteins, which showed a close relationship with protease or meat quality traits in PM muscle, did not show a strong relationship with protease or meat quality traits in IL muscle. For IL muscle, glceraldehyde-3-phosphate dehydrogenase and ATP synthase subunit beta were linked to CIE L* and pH, while cytochrome c oxidase polypeptide VIIc, glucose-6-phosphate isomerase, and myosin light chain 1 were linked to CIE b*. In addition, L-lactate dehydrogenase, cytoplasmic actin, ADP/ATP translocase, and myosin binding protein C1 were associated with cooking loss, whereas myoglobin, peptidyl-prolyl cis-trans isomerase, myosin motor domain-containing protein were closely related to calpain. Hemoglobin subunit beta, adenylate kinase isoenzyme 1, LIM domain binding 3, and uniquinol-cytochrome c reductase core protein 2 were linked to caspase-3 and proteasome 20S, while actin alpha 1 and creatine kinase were linked to cathepsin B.
Discussion
Meat quality during storage is influenced by external and internal factors. In particular, intrinsic factors, such as muscle fiber type and proteolytic enzyme activity, determine postmortem changes in meat quality characteristics. Poultry meat is generally classified as white meat due to the high composition of white muscle fiber (type IIB; Joo et al., 2013). Similar to the findings of Huo et al. (2021), duck PM has only fast-type (type II) fibers in the present study. However, another previous study (Kim et al., 2008), which revealed that duck PM consisted of 73.3% type IIB and 26.7% type IIA muscle fibers, is inconsistent with the present study. That is, duck PM muscle fibers can be classified into two types (IIA and IIB) according to their contractile or metabolic properties, whereas muscle fibers can be distinguished by the distribution of myosin isoforms (slow and fast; Kim et al., 2013). Duck IL muscle is composed of both slow and fast types of muscle fiber due to the different morphological and physiological properties from those of PM. The different distribution of muscle fiber types results in the differences in meat quality characteristics between PM and IL muscles.
The relationships between protein hydrolysis of endogenous enzymes and meat quality characteristics, especially meat tenderization, have been demonstrated (Bekhit et al., 2014; Huang et al., 2011). In addition, Chéret et al. (2007) reported that cathepsin L plays a role in the tenderization process of bovine and fish muscles, while cathepsin B plays a role only in that of fish muscles. A similar role was found in the present study in which the activity of cathepsin L changed after 10 days of storage, but cathepsin B showed no difference in its activity. Calpains have been proven to be the main enzyme in the meat tenderization process in most species (Lana and Zolla, 2016). In the present study, a significant decrease in calpain activity was detected in PM and IL muscles of duck meat from the first day to the tenth day. This was supported by the result of the degradation of myofibrillar proteins such as actin, MHC, and troponins. Robert et al. (1999) used incubation experiments to demonstrate that proteasome 20S can effectively degrade bovine myofibrils and interfere with the role of proteasome 20S in postmortem proteolysis. However, in the present study, proteasome 20S activity was not different in IL muscle throughout the storage period, while in PM muscle, its activity decreased after 10 days of storage. Caspase-3 activity was decreased in PM muscle during cold storage. In the present study, the activities of proteases were maintained or gradually decreased from the initial storage time to the later storage stages, but tenderness was not affected by the proteases, as observed previously (He et al., 2019).
Several proteins showed the relationships between their degradation trends and protease activity or meat quality traits, especially meat color and water-holding capacity. In particular, meat color traits were majorly related with the degradation of enzymes responsible for glycolysis, such as glycerol-3-phosphate dehydrogenase, phosphoglucomutase 1, glyceraldehyde-3-phosphate dehydrogenase, and glucose-6-phosphate isomerase as reported in previous study which demonstrate the correlations among glycolytic dehydrogenase including glyceraldehyde-3-phosphate dehydrogenase, lactic dehydrogenase, and meat color stability were observed in ovine muscle (Xin et al., 2018). For water-holding capacity, Joo et al. (1999) demonstrated that the rate of drip loss declined according to the increasing of the sarcoplasmic protein solubility. In the present study, cooking loss in PM muscle was closely related to the degradation of proteins, such as fructose-bisphosphate aldolase, phosphagen kinase N-terminal domain-containing protein, and malate dehydrogenase, and these proteins were also associated with the activities of cathepsin B and calpain. However, IL muscle showed different trends in the relationship between protein and water-holding capacity, as observed in the relationship between cooking loss and myosin binding protein C1, cytoplasmic actin, ADP/ATP translocase, and L-lactate dehydrogenase. This was supported in part by a previous study (Hamm, 1986), which showed that myosin and actin had a significant effect on water-holding capacity of meat.
Conclusion
The differences in muscle fiber distribution between duck PM (fast type) and IL (slow and fast) resulted in different trends in proteolysis and changes in meat quality during cold storage. Large amounts of various proteolysis-induced peptides were observed in both muscles at the initial storage time (day 1), and this indicates that intrinsic proteolytic enzymes play a major role in protein degradation at the early postmortem stage. The proteins responsible for glycolysis or myofibrillar proteins were closely related to changes in meat color or water-holding capacity during cold storage. These results indicate that the proteolysis is affected by storage time and affects the change in duck meat quality during storage. This phenomenon also depends on the distribution of muscle fiber types, which is determined by physiological properties of the muscle.