










Introduction
Meat tenderness is generally considered by the consumer to be the most important palatability factor. Post-mortem tenderization in skeletal muscle is a complex phenomenon that has yet to be fully understood. Muscle is tender right after the animal is slaughtered but once the onset of rigor mortis, a permanent cross-link between actin and myosin proteins is formed and this leads to muscle toughness (Bowker et al., 2010). Beef requires ageing for at least 14 d in a controlled environment with temperature ranging from −1°C to 5°C to achieve tenderness (Christian and Stephen, 2010). During this period, the degradation of muscle contributes to meat tenderness. This process is known as post-mortem aging (also known as maturation or conditioning) of meat and is widely practiced by beef producers (Chéret et al., 2007; Christian and Stephen, 2010).
Sarcoplasmic proteases are crucial in protein catabolism and post-mortem muscle softening. It is believed that two main proteolytic systems are involved in the tenderization of meat: the cathepsins and the calpains. Many researchers consider the calpain system to be the major contributor to meat tenderness during post-mortem aging (Koohmarie and Geesink, 2006). However, this assumption has been debated (Herrera-Mendez et al., 2006) and the role of cathepsins is not fully understood yet. Cathepsins exhibit greater heat stability than calpains (Laakkonen et al., 1970; Pomponio and Ertbjerg, 2012) as the latter have been reported to completely inactivate at temperatures above 55°C whereas cathepsins, particularly B and L have been reported to remain active even after 24 h of heating at 55°C (Christensen et al., 2011; Ertbjerg et al., 2012). Cathepsin D has also been reported to have a lower heat resistance compared with cathepsin B and L (Spanier et al., 1990). Cathepsin B and L are endopeptidases that may contribute to meat tenderness by weakening collagen in connective tissue, leading to its increased solubility (Agarwal, 1990; Christensen et al., 2013; Solvig, 2014). Incubation of connective tissue with cathepsin B has been reported to significantly decrease the denaturation temperature of connective tissue from both calf and steer (Beltrán et al., 1992). In addition, a study by Burleigh et al. (1974) has shown that cathepsin B contributes to the degradation of both soluble and insoluble collagen by eliminating intermolecular cross-links.
In addition to proteolysis during ageing, meat tenderness may continue to develop during the cooking step, particularly when the meat is heated for a longer duration at temperature that is optimum for enzyme activity (Ertbjerg et al., 2012). Sous vide is a method of cooking vacuum-packaged food at a precise temperature for a long duration, from hours to days (Baldwin, 2012). This form of cooking helps to retain moisture and is known to produce tender and juicy meat (Laakkonen et al., 1970). Temperatures from 55°C to 80°C and cooking for 6 to 48 h are typical conditions for cooking meat (Baldwin, 2012).
We hypothesized that cathepsins would remain active during sous vide cooking and could still contribute to the tenderization process of beef brisket during the cooking process as opposed to cooking meat at high heat such as grilling whereby high cooking temperature and time is often associated with the toughening of meat. Limited studies have been done to examine the effects of sous vide cooking on the proteolytic enzyme activities in tough beef muscle cuts such as brisket. Thus this investigation was designed to study the effects of (1) post-mortem storage and (2) sous vide cooking at different temperatures (50°C−70°C) for up to 24 h on the activities of the B, L, and H cathepsins in beef brisket, which will aid in determining the contribution of these cathepsins to tenderness of tough cuts of meat.
Materials and Methods
Hot-boned briskets from three steers at 4 h post-mortem were kindly provided by a local slaughterhouse (ANZCO Foods, Bulls, New Zealand) and immediately transported to the laboratory. At approximately 6 h post-mortem, muscles were cut into small samples of 2 cm3 or thin strips after removal of the visible subcutaneous fat, vacuum packed into portions and stored in either 4°C or −20°C. Post-mortem storage and sous vide experiments were two separate experiments that were run in parallel, with two different objectives, but with the samples from the same carcasses. Thus, for each carcass, the samples were divided into three batches as described in Fig. 1: as control, and for post-mortem storage and sous vide experiments. Three samples per treatment from different carcasses were analysed for pH and cathepsin B and H activities as described in sections 2.5 and 2.6.

The experiment to assess the effects of post-mortem storage conditions on cathepsin activities was divided into short-term (6 h to 4 d post-mortem at either 4°C or −20°C) and long-term storage (further 4–18 d storage at either 4°C or −20°C of the 4 d refrigerated meat).
The vacuum-packed muscle samples were stored in a chiller or freezer at 4°C and −20°C, respectively. At the end of the allocated storage time (Fig. 1), samples were assayed for pH and cathepsin activities.
One-day post-mortem meat stored at 4°C was chosen for performing the sous vide experiments, based on previous reports (Chéret et al., 2007). Slight modifications were made to the sample preparation procedures reported by Ertbjerg et al. (2012) Meat was cut into small strips, vacuum packed and stored at 4°C on the day of slaughter (Fig. 1). At one day post-mortem, the vacuum bags containing the meat strips were cooked in water baths set at 50°C, 55°C, 60°C, 65°C, and 70°C for 1, 5, or 24 h. At the respective time interval, samples were removed from the water bath and cooled in ice water to below 25°C. The temperature of the water bath was monitored using a digital thermometer (Q1437, Dick Smith Electronics, Chullora, Australia) to ensure that the desired temperature had been reached before fully submerging the bags into the water bath. Since the samples were cut into thin strips and as confirmed from previous experimentation (Zhu et al., 2018), it was assumed that the core temperature of the samples reached the water bath temperature quite quickly (in less than an hour).
Three random cooked samples from each carcass were homogenised together and assayed for pH and cathepsin activities. All samples were stored at −20°C until analyzed. Cathepsin activities were calculated for both cooked and raw samples and expressed as a relative activity (%):
where At and A0 are the enzyme activities of cooked and raw samples, respectively.
The pH of the meat homogenates were determined with a glass electrode pH meter (Cyberscan pH 510, Eutech Instruments, Vernon Hills, IL, USA). The pH meter was calibrated using pH 7.0 and 4.0 standard buffers stored at room temperature. Meat homogenate was prepared by blending finely-chopped meat with milli-Q water in a ratio of 1:10 for 1 min using a food processor (BFP100WHT, Breville, Sydney, Australia).
Sarcoplasmic protein extract was prepared using the method described by Chéret et al. (2007) with slight modifications. Three random muscle samples from a single carcass were finely chopped and homogenized with an extraction buffer comprising of Tris–HCl, 2-mercaptoethanol and ethylenediaminetetraacetic acid in a ratio of 1:3 for 1 min using a food processor (BFP100WHT, Breville). The homogenized mixture was centrifuged at 25,000×g for 20 min at 4°C in a Sorvall Evolution RC centrifuge (Thermo Fisher Scientific, Waltham, MA, USA). The supernatant was collected and filtered using 0.45 μm syringe filter and referred to as crude extract. This crude extract was immediately used for the cathepsin assays.
Activities of cathepsin B, H, and L were analyzed in the sarcoplasmic extract using the method described by Chéret et al. (2007). The cathepsin activities were determined at room temperature in a 96-well microplate, consisting of 6 μL of 5% CHAPS prepared in milli-Q water; 1 μL of 1.40 M 2-mercaptoethanol; 16 μL of 5% (w/v) Brij® 35 prepared in milli-Q water; 5 μL of 20 mM synthetic fluorogenic substrate prepared in methanol and 70 μL of 0.4 mM acetate/acid acetic (pH 4) buffer containing 10 mM 2-mercaptoethanol and 1 mM EDTA. The substrates for cathepsin B, cathepsin B and L, and cathepsin H were Z-Arg-Arg-7-amido-4-methylcoumarin hydrochloride (C5429, Sigma-Aldrich, St. Louis, MO, USA), Z-Phe-Arg-7-amido-4-methylcoumarin hydrochloride (C9521, Sigma-Aldrich), and L-Arginine-7-amido-4-methylcoumarin hydrochloride (A2027, Sigma-Aldrich), respectively. The reaction was initiated by the addition of 200 μL of crude sarcoplasmic protein extract. The fluorescence intensity was determined using a microplate reader (Wallace Victor2 1420 multilabel counter, Perkin Elmer, Waltham, MA, USA) with excitation and emission wavelengths of 355 nm and 460 nm, respectively. A control was run in parallel in which the protein extract was substituted by extraction buffer. Cathepsin specific activities were expressed in FU (units of fluorescence) increase per min per g of muscle. Cathepsin L activity was calculated by subtracting cathepsin B from the cathepsin B+L activity. Cathepsin D activity could not be measured in the samples due to technical issues in standardization of cathepsin D assay. The difficulties in standardising cathepsin D assay may be attributed by its low activity level in meat and meat products as reported by Rico et al. (1991a). Hence, this paper does not report cathepsin D activities in the samples.
Significant differences in pH and enzyme activities among different treatments were determined by one-way ANOVA Tukey test at the 95% significance level using Minitab® 17 (Minitab, 2014).
Results and Discussion
There was no significant difference in pH value and cathepsin H activities during 4 d of ageing at both chilled and frozen storage conditions (Table 1). Although there was no significant difference in cathepsin H activities, the numerical difference between the control and frozen samples could be because L-Arginine-7-amido-4-methylcoumarin (substrate for cathepsin H) was also cleaved by aminopeptidase, and under the conditions of the assay some residual peptidase activity may be present (Toldrá and Etherington, 1988).
For cathepsin B, there was a significant increase (compared to 6 h post-mortem) in its activity after 4 d of ageing at both temperatures however no significant difference was observed among the storage temperatures (Table 1). Similar increase for cathepsin B was observed for sea bream muscles, which was attributed to enzyme activation by low pH (Matos, 2013). Cathepsin B has an optimum pH of 5.5 towards most substrates, which is near the ultimate pH of the meat. During post-mortem storage when the temperature and pH decrease, the fragile membranes of lysosomes may rupture resulting in the release of cathepsins (Bowker et al., 2010; Lana and Zolla, 2016). Despite the fact that no significant difference was observed in the pH during 4 d of ageing, it is possible that the decrease in temperature during storage or the formation of ice crystals may have ruptured the lysosomes, releasing cathepsins, therefore contributing to an increase in cathepsin B activity.
There was no significant change in pH value throughout subsequent 2 wk’s storage at both temperatures (Table 2). The activities of cathepsins B, L, and H also remained stable and unchanged during this storage period. Similar results have been reported for ostrich fillet where cathepsin B and L showed no decrease in their activities after 12 d storage at 2°C to 4°C (van Jaardveld et al., 1997). Previous studies have indicated that all cathepsins are capable of degrading myosin (Allen and Goll, 2003) but no myosin degradation has been reported during post-mortem storage at 0°C to 4°C (Bandman and Zdanis, 1988). Moreover, the changes in shear force values during post-mortem ageing for 2 wk at 1.2°C for three types of bovine muscles were found to be different despite having similar level of cathepsins B, H, and L (Koohmaraie et al., 1988). Thus, the tenderizing effect of cathepsins during the long term storage at refrigerated temperatures remains questionable. Other proteolytic systems such as caspase, metalloproteases, thrombin, and plasmin may also be involved during post-mortem ageing (Ouali et al., 2013).
At all temperatures, increasing the cooking time did not have a significant effect on the pH (Table 3). There was a significant increase (p<0.05) in pH when the temperature increased from 50°C to 70°C, after 1 and 5 h of cooking. An increase in pH during cooking has also been observed for bovine muscles (longissimus, semitendinosus, and rectus femoris) heated at 60°C for 10 h (Laakkonen et al., 1970). Small increments in pH upon cooking of meat at 60°C have been reported to be due to a decrease in acidic groups in the meat proteins (Hamm and Deatherage, 1960).
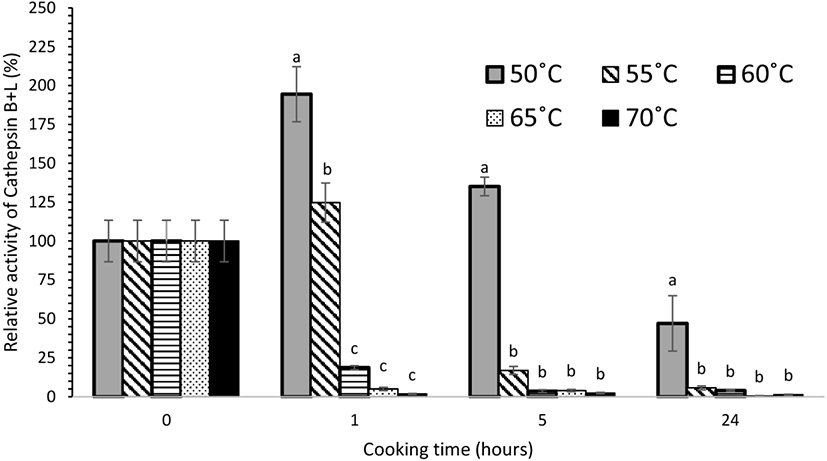
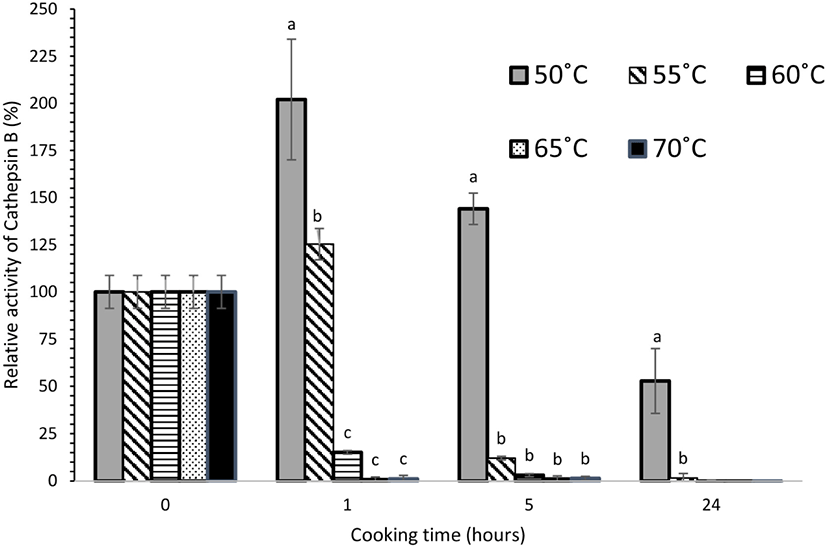
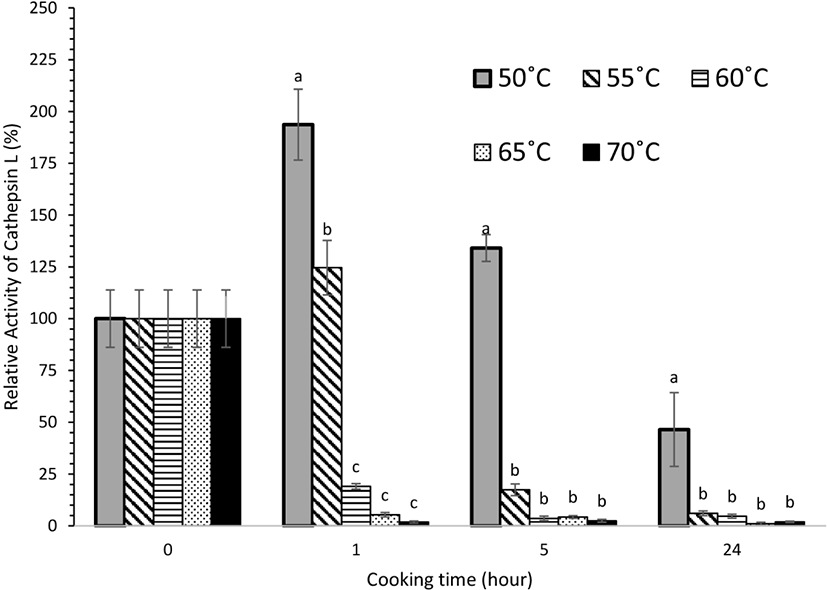
Cathepsin B+L activity was heat stable and these proteases remained active throughout cooking at 50°C even after 24 h and for the first 5 h at 55°C (Fig. 2). At 50°C, the cathepsin B+L activity increased significantly (p<0.05) after 1 h, while further cooking led to a decrease in activity. There was a significant reduction (p<0.05) in cathepsin B+L activity after cooking at 60°C, 65°C, and 70°C, where most of the extractable activity was lost after 1 h. Subsequently no significant change (p>0.05) in cathepsin B+L activity was observed for meat cooked from 5 to 24 h at these temperatures. The reduction in activity is likely to be due to the heat sensitivity of cathepsin B and L, combined with the effect of the increase in pH observed for these treatment conditions. A similar trend was observed for cathepsin B (Fig. 3) and cathepsin L (Fig. 4) activity. In a study on porcine longissimus muscle, cathepsin B+L activity was reported to increase with an increase in temperature from 48°C to 58°C (Christensen et al., 2011). In another study conducted on beef semitendinosus muscle, the activity of cathepsins B and L in the expelled cooking loss was highest after cooking at 53°C for 2.5 h and then decreased with increasing temperature and time (Christensen et al., 2013). All these observations are consistent with our experimental results. In this present study, no significant cathepsin B+L activity was detected after 5 h heating at 60°C. However, a higher heat tolerance of cathepsins B and L has been reported for beef semitendinosus muscle where their activity was measurable even after 19.5 h at 63°C (Christensen et al., 2013). This could be because of the differences among the meat cuts used in both the studies. Ertbjerg et al. (2012) found that cathepsin B+L activity reached a maximum after heating at 55°C for 1.5 h in porcine longissimus muscle. They suggested that part of cathepsin B and/or cathepsin L may exist in the form of a pro-enzyme which is activated by heat. An increase in activity was also observed in our experimental results, which was evident after 1 h of heating at 50°C. Increases in collagen solubilization and tenderness were also evidenced in this temperature range (Christensen et al., 2011). The synergistic effect of heat denaturation and proteolytic action of cathepsins (B and L) has been reported to account for an increased weakening effect on collagen that led to more tender meat during sous vide cooking at temperatures <55°C (Dominguez-Hernandez et al., 2018).
At all temperatures, a significant reduction (p<0.05) of cathepsin H activity occurred during the first hour of cooking (data not shown). At 50°C, cathepsin H remained active (15% of initial activity) during the first hour but it lost most of the extractable activity within 5 h. At 55°C and above, no extractable activity was detected after 1 h of cooking. In addition, there was no significant difference (p>0.05) in cathepsin H activity after 1 h of cooking at temperature ranging from 55°C to 70°C. Thus, it is unlikely that cathepsin H is responsible for the tenderization effect usually observed during sous vide cooking of meat at temperatures <70°C.
Conclusion
During post-mortem storage, only cathepsin B activity was observed to increase from 6 h to 4 d post-mortem at both (refrigerated and frozen) storage conditions. There were no significant changes in cathepsin B, H, and L activities during long-term storage of two weeks. For the sous vide experiments, the increase in cathepsin B+L activity at 50°C after 1 h of cooking suggests that cathepsin B and/or L in beef brisket may also exist in the form of a pro-enzyme, which is activated by heat. Thus, at this temperature, with a higher cathepsin B+L activity, these enzymes are likely to be involved in proteolysis and contribute to the tenderizing effect. Cathepsin B and L were found to be more heat stable at sous vide temperatures (50°C for 24 h, 55°C for 5 h, and 60°C and 70°C for 1 h) compared to cathepsin H, supporting the hypothesis that cathepsin B and L remain active at typical sous vide cooking temperatures and could be involved in the tenderization process.