










Introduction
Skeletal muscle is generally composed of several types of fiber, which are subdivided by differences in morphological, contractile, and metabolic properties (Kim et al., 2014; Pette and Staron, 2000). Several methods have been used to classify muscle fiber types. The myosin ATPase histology, which is based on the sensitivity of each myosin ATPase on myosin heavy chains (MHC) to pH pre-incubation (Brooke and Kaiser, 1970; Guth and Samaha, 1969). According to the different activities of metabolic enzymes, such as succinate dehydrogenase and glycerol-3-phosphate dehydrogenase, two or more types of muscle fiber can be subdivided (Blanco et al., 1988; Martin et al., 1985). However, these methods, which are dependent on the enzyme activities, require biopsies or samples taken promptly from postslaughtered specimens. Moreover, they have disadvantages for distinguishing hybrid fiber types. Immunohistochemistry using several antibodies specific to MHC isoforms allows us to identify hybrid fibers (Kim et al., 2014). However, this method requires multiple cross-sections. Despite these disadvantages and inconveniences for muscle fiber typing, these methods have been widely used because muscle fiber characteristics are closely related to carcass traits and meat quality, as previously reported in the literatures (Chang et al., 2003; Choi et al., 2007; Eggert et al., 2002; Gil et al., 2001; Gonzalez et al., 2008; Hwang et al., 2010; Larzul et al., 1997; Micol et al., 2009; Ozawa et al., 2000; Ryu et al., 2008; Wojtysiak and Poltowicz, 2014).
Bloemberg and Quadrilatero (2012) suggested the improved method for rapid identification of MHC-based muscle fiber types in rat, mouse, and human skeletal muscles using immunofluorescence analysis. This procedure is time efficient, because primary antibodies specific to each MHC isoform and secondary antibodies can be used as a cocktail. Furthermore, this method allows to easily identify hybrid types as well as pure types of muscle fiber. To adopt this useful procedure for muscle fiber typing in livestock animals such as pigs and cattle, some modifications are required. Although commercial antibodies, which are specific to MHC isoforms, are available, few antibodies are species-specific as summarized by Quiroz-Rothe and Rivero (2004).
Therefore, the objective of this study was to optimize the method for rapid and efficient muscle fiber typing in skeletal muscles of livestock animals such as cattle and pigs. Among several antibodies that are specific to MHC isoforms, we selected the appropriate antibodies against MHC isoforms of bovine and porcine skeletal muscles, and investigated rapid procedures for identification of pure and hybrid fiber types in bovine and porcine skeletal muscles.
Materials and Methods
Four major muscles such as longissimus thoracis (LT), psoas major (PM), semimembranosus (SM), and semitendinosus (ST), were obtained from pigs (n=10, barrows, 73.5±2.4 kg of carcass weight) and cattle (n=10, steers, 437.2±7.8 kg of carcass weight), respectively. All muscles were taken from the left side of each carcass at 24 h postmortem at the commercial slaughterhouse. Three pieces (1.0×1.0×1.5 cm) of muscle specimens were cut from each muscle, promptly frozen in 2-methylbutane chilled with liquid nitrogen, and kept at −80°C until analysis.
Transverse sections were cut into 10 μm thickness using a cryostat (CM1520, Leica Biosystems, Wetzlar, Germany) at −20°C. The sections were pre-incubated in 10% normal goat serum (Cell Signaling Technology, Danvers, MA, USA) for blocking. Primary antibodies (DSHB, IA, USA) were used for detecting one or more MHC isoforms (BA-F8, slow/I; SC-71, 2a and 2x; BF-35, all isoforms except for 2x; 6H1, 2x; BF-F3, 2b). For multicolor immunofluorescence, Alexa Fluor 350, 488, 594, and 647 (Thermo Fisher Scientific, Waltham, MA, USA) were applied to each section for 1 h at room temperature. Primary and secondary antibodies were applied serially or in a cocktail to the sections, respectively. The dilution ratio and configuration of antibodies are presented in Table 1. All the sections were rinsed in PBS for 5 min with triplication after incubation. The sections were visualized with confocal scanning laser microscope (TCS SP8 STED, Leica Biosystems, Wetzlar, Germany). Cross-sectional area (μm2), relative number composition (%), and relative area composition (%) of each muscle fiber type were analyzed from approximately 800 fibers per section using Image Pro Plus program (Media Cybernetics, Rockville, MD, USA).
As shown in Table 2, muscle fiber types were classified according to the distribution of MHC isoforms. In bovine skeletal muscles, three pure types, I, IIA, and IIX, were identified by BA-F8 (anti-MHC slow/I), SC-71 (anti-MHC 2a and 2x), BF-35 (anti-MHC slow/I and 2a), and 6H1 (anti-MHC 2x). Hybrid fiber type IIA+IIX was identified by SC-71, BF-35, and 6H1. Muscle fibers in porcine muscles were classified into seven fiber types including four pure types (I, IIA, IIX, and IIB) and two hybrid types (IIA+IIX and IIX+IIB). Each pure type was identified by BA-F8, SC-71, BF-35, and BF-F3, whereas hybrid types were identified by a combination of SC-71 and BF-35 for IIA+IIX and by a combination of SC-71 and BF-F3 for IIX+IIB.
All data are presented as means±SE. The general linear model (GLM) was adopted to analyze the differences in muscle fiber characteristics (cross-sectional area, relative number composition, and relative area composition) among the muscle fiber types within each muscle (LT, PM, SM, and ST) in cattle and pigs. Duncan's multiple range test was used to determine the significant differences in muscle fiber characteristics using the statistical analysis software (SAS Institute, Cary, NC, USA) at a 5% level of significance.
Result and Discussion
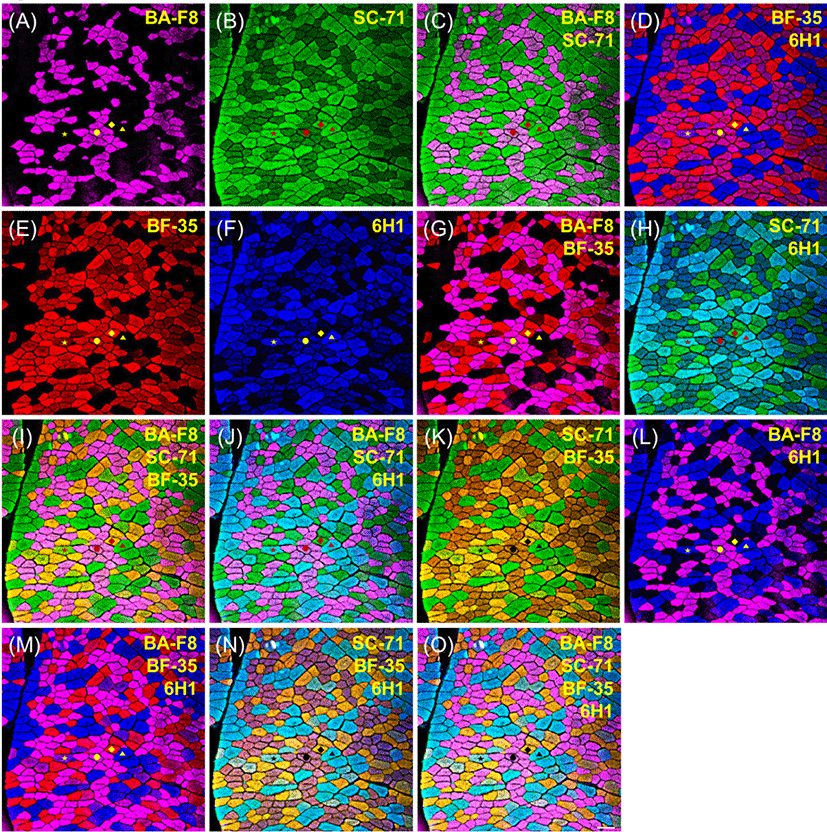
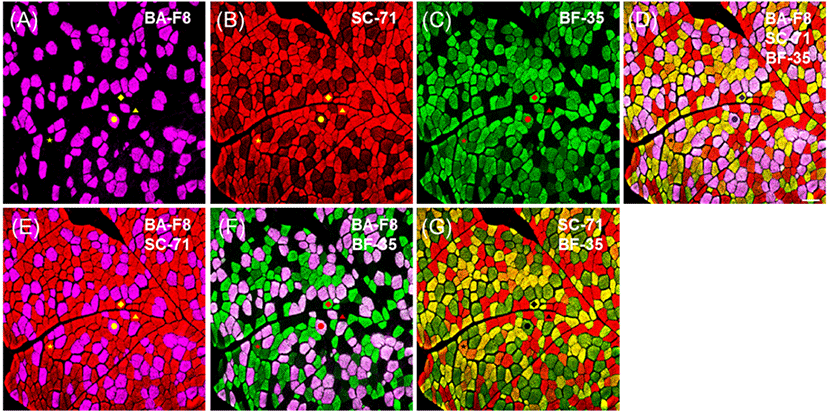
Representative photomicrographs of bovine muscle reacted with four monoclonal antibodies are presented in Fig. 1. Three pure types, I, IIA, and IIX, were identified by a combination of four anti-MHCs, including BA-F8, SC-71, BF-35, and 6H1, as shown in Table 3. Previously, those antibodies had been adopted to identify muscle fiber types regardless of species (Table 2). Muscle fiber type IIB was not detected by any anti-MHC antibodies in the major bovine muscles (LT, PM, SM, and ST), as is consistent with previous studies: MHC 2b isoform (corresponding to type IIB) was not detected at the gene (myosin 4) as well as protein levels in skeletal muscles of adult cows and steers (Chikuni et al., 2004;Kim et al., 2016). Three pure types were easily identified by the combination of two anti-MHC antibodies: BA-F8 and BF-35 (Fig. 1G); SC-71 and 6H1 (Fig. 1H); SC-71 and BF-35 (Fig. 1K); BA-F8 and 6H1 (Fig. 1L). In addition, combinations of three or more anti-MHC antibodies can be used to identify three pure types, as shown in Fig. 1 (I, J, M, N, and O). Hybrid fiber types such as I+IIA and IIA+IIX could be expected from the bovine muscles. Hybrid type I+IIA can be identified by a combination of BA-F8, SC-71, and 6H1 (Fig. 1J); however, this type was not found in the present study (Fig. 1). Another hybrid fiber type, IIA+IIX, was detected by a combination of BF-35 and 6H1 (Fig. 1D). Additionally, the combinations with three antibodies, such as a set of BA-F8, BF-35, and 6H1 (Fig. 1M) or a set of SC-71, BF-35, and 6H1 (Fig. 1N), allowed to detect hybrid fiber type IIA+IIX. Although hybrid fibers were confirmed with those combinations, their fluorescence intensities were lower than that of pure types, as observed by Bloemberg and Quadrilatero (2012). To improve the resolution of the hybrid type, secondary antibodies conjugated with other fluorescent dyes were used and the visualized sections are presented in Fig. 2. A combination of three anti-MHC antibodies such as BA-F8, SC-71, and BF-35 was adopted, and Alexa Fluor 594 and 488 were used for detection of SC-71 and BF-35, respectively. Consequently, the color of hybrid type IIA+IIX can be distinguished from those of the fiber types I, IIA, and IIX (Fig. 2D and 2G).
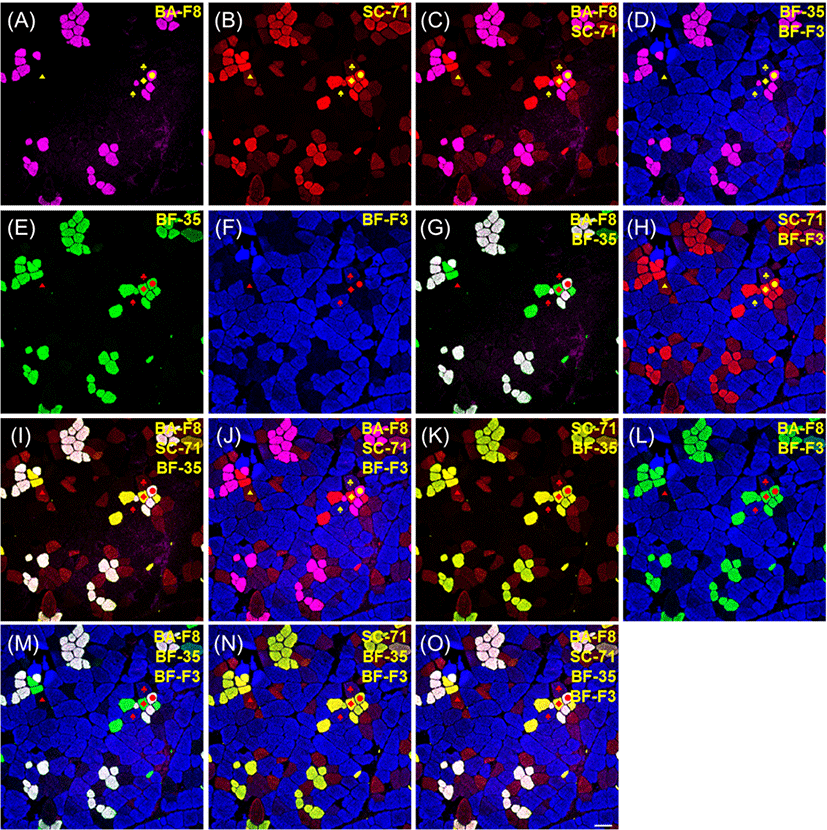
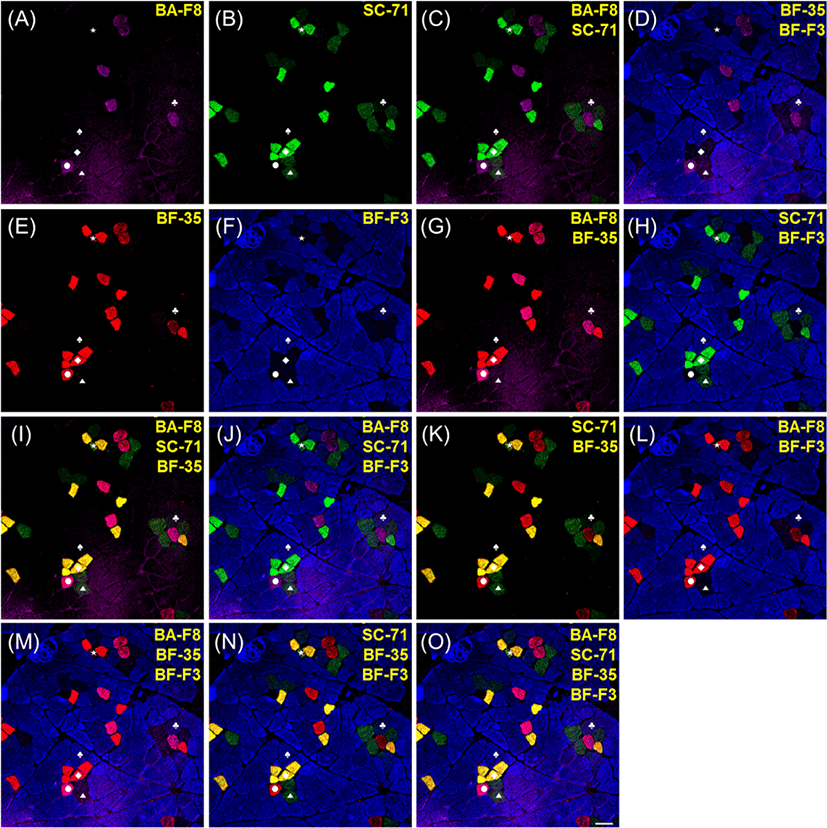
The results of immunofluorescence of porcine muscle, which were reacted by four monoclonal antibodies, such as BA-F8, SC-71, BF-35, and BF-F3, are presented in Figs. 3 and 4. Muscle fiber type IIX in bovine muscle was identified by 6HI antibody; however, this antibody did not work in any porcine muscles. Thus, 6H1 was not adopted for muscle fiber typing of porcine muscles. Among the four anti-MHC antibodies, BA-F8, BF-35, and BF-F3 showed the same specificity to MHCs as previously observed in porcine muscles (Kim et al., 2014; Lefaucheur et al., 2002; Quiroz-Rothe and Rivero, 2004). However, SC-71 had a different reactivity to MHCs different from that in the previous studies. In porcine skeletal muscles, SC-71 generally reacted with MHCs IIA and IIX with different intensities (Kim et al., 2013; Lefaucheur et al., 2002). In the present study, SC-71 reacted with MHC I as well as MHCs IIA and IIX (Fig. 3B). Thus, hybrid fiber type I+IIA could not be identified. From the sections with serial staining procedure, the normal reactivity of SC-71 was observed to have strong intensity with type IIA and weak intensity with type IIX (Fig. 4B). Four pure types (I, IIA, IIX, and IIB) could be detected by combinations of two or more anti-MHC antibodies: BA-F8 and SC-71 (Figs. 3C and 4C); BA-F8, SC-71, and BF-35 (Figs. 3I and 4I); BA-F8, SC-71, and BF-F3 (Figs. 3J and 4J); BA-F8, BF-35, and BF-F3 (Figs. 3M and 4M); BA-F8, SC-71, BF-35, and BF-F3 (Figs. 3O and 4O). The expectable hybrid fiber types were I+IIA, IIA+IIX, and IIX+IIB; however, IIA+IIX and IIX+IIB were detected in the present study. Hybrid type IIA+IIX can be distinguished by a combination of SC-71 and BF-35, whereas type IIX+IIB can be identified with a combination of SC-71 and BF-F3. Among the hybrid fiber types, those two types can be confirmed using both cocktail and serial staining procedures. Thus, our findings indicate that hybrid fiber types IIA+IIX and IIX+IIB can be identified by both cocktail and serial staining procedures, but hybrid fiber type I+IIA can be detected not by cocktail but by serial staining procedure. In other words, the method of cocktail staining could not be recommended for porcine muscle fiber typing despite its time efficiency. To help to distinguish each muscle fiber type, secondary antibodies replaced with other antibodies conjugated with different fluorescent dyes (Alexa Fluor 594 to 488 for SC-71 and 488 to 594 for BF-35) consequently improved resolutions for type I and IIA (Fig. 4O).
Muscle fiber size (CSA, μm2) and composition (relative area and relative number, %) was investigated in bovine and porcine skeletal muscles are presented in Table 4. The improved procedures for immunofluorescence by combination of anti-MHC antibodies and fluorescent dyes allowed us to identify four (three pure and one hybrid) fiber types in bovine muscle and six (four pure and two hybrid) fiber types in porcine skeletal muscle. In bovine muscles, hybrid type IIA+IIX was detected with ranges from 1.90% to 4.16% of relative numbers and from 1.97% to 5.16% of relative area. Previously, three fiber types, I, IIA, and IIX (or IIB), were observed in bovine muscles (Hwang et al., 2010; Kim et al., 2016; Ozawa et al., 2000), whereas hybrid fiber types were not frequently identified (Moreno-Sanchez et al., 2008; Greenwood et al., 2009). Fiber type IIB and hybrid type IIX+IIB were not detected in bovine muscle because of the lack of MHC 2b in major bovine muscles (LT, PM, and SM), as previously reported (Kim, 2014; Kim et al., 2016). Muscle fiber composition showed a similar trend to the results of previous reports (Hwang et al., 2010; Ozawa et al., 2000). However, muscle fiber size of LT muscle was not consistent with previous findings. There was no significant (p>0.05) difference in CSA of all fiber types in LT muscle.
1 Muscle fiber types were identified by monoclonal antibodies being specific to myosin heavy chains, as shown in Table 3.
In porcine skeletal muscles, except for PM, hybrid fiber types IIA+IIX and IIX+IIB were identified; they were distributed from 3.63% to 5.99% of relative numbers and from 4.83% to 7.56% of relative area. It has been reported that porcine muscles are composed of 1.7%–11.9% of hybrid fiber types IIA+IIX and IIX+IIB (Kim et al., 2013). The other hybrid type, I+IIA, was previously reported as being observed in rodent, bovine, and canine skeletal muscles (Acevedo and Rivero, 2006; Moreno-Sanchez et al., 2008). However, there was no hybrid type I+IIA in the present study, perhaps because these were different species. Muscle fiber size showed significant differences between type I and type IIB, and type IIB was larger than type I regardless of muscle types (p>0.05). Fiber size of hybrid type IIX+IIB was larger than that of another hybrid type, IIA+IIX, in porcine LT muscle, as is consistent with our previous report (Kim et al., 2014). Unlike LT muscle, SM showed an opposite trend in hybrid fiber size: type IIA+IIX (6,631 μm2 of CSA) was bigger than type IIX+IIB (4,520 μm2 of CSA). The result of muscle fiber compositions (relative area and relative number) of porcine muscles except for ST was consistent with previous studies (Kim et al., 2014; Ryu et al., 2019) showing that relative area or number of type IIB was higher than those of type I, whereas ST was observed to have the same amount of type I as of type IIB. This finding indicates that muscle fiber compositions can be dependent on muscle type.
Conclusion
To optimize typing of muscle fiber in bovine muscle, the combinations of three or four anti-MHC antibodies (BA-F8, SC-71, BF-35, and 6H1) are required, and the cocktail staining procedure with primary and secondary antibodies could save time. Porcine muscle fibers could be typed using four anti-MHC antibodies (BA-F8, SC-71, BF-35, and BF-F3). Unlike for the bovine muscle, cocktail procedure is not recommended in porcine muscle, because of the abnormal reactivity of SC-71. For improved resolution, secondary antibodies conjugated with different fluorescent dyes allowed us to distinguish individual types of muscle fiber in bovine as well as porcine muscles. Accordingly, four muscle fiber types, including one hybrid type, were identified in bovine muscles, and six fiber types (four pure types and two hybrid types) were detected in porcine muscles. However, the hybrid fiber type I+IIA was not found in either species. Differentially distributed muscle fiber types were dependent on species and muscle type.