










Introduction
Meat is highly perishable during storage because of its high water content and nutrients, such as proteins, lipids, vitamins, and minerals, which are suitable for microbial growth (Amit et al., 2017; Kim et al., 2019). Globally, 20% of meat produced is lost or wasted every year. The spoilage of meat by microorganisms during storage is one of the major reasons for this (FAO, 2011). Therefore, improving meat shelf life by inhibiting microbial growth is a potential food security strategy because the quantitative growth of livestock is limited (Alexander et al., 2017).
Freezing allows long-term storage of food by decreasing the temperature and water activity (Leygonie et al., 2012). However, freezing changes the functional properties of muscle proteins (Lee et al., 2021a). The functional properties of meat proteins, such as the interaction between protein and protein, protein and water, and protein and fat, are very important factors in determining the quality of comminuted meat products (Sun and Holley, 2011). Since the free energy barrier for protein unfolding is decreased at the ice-water interface under low temperatures, structural modification occurs during freeze-storage (Arsiccio et al., 2020). Moreover, proteins can be denatured by increasing the solute concentration in unfrozen water (Estévez et al., 2011). Therefore, denatured proteins in thawed meat can exhibit deteriorated processing characteristics. Previous studies have reported reduced gelling and emulsifying capacities of meat proteins by the denaturation of myofibrillar proteins after the freezing-thawing cycle (Chan et al., 2011; Xia et al., 2010). However, freeze-thawed pork is generally used to produce meat products in processed meat industries because the deteriorated functionality of proteins can be improved with the use of salts, such as sodium chloride and phosphate (Ruusunen and Puolanne, 2005).
In addition, drying has many economic advantages for freezing. These include convenient transportation and increased storage efficiency owing to the decreased volume and weight of the meat. Additionally, dried foods packaged with non-moisture-permeable materials can be stored at room temperature (Amit et al., 2017); thus, energy does not need to be used for cooling. Freeze-drying or lyophilization is a conventional technique used in both the food and pharmaceutical industries to improve the storage and distribution of proteins with high stability (Ratti, 2001; Roy and Gupta, 2004). Nevertheless, changes in protein functionality with either reversible or irreversible conformational changes induced by dehydration have been reported because water is an important component of the native protein structure. Moreover, hydrogen bond interactions with water molecules are lost during protein dehydration (Griebenow and Klibanov, 1995; Prestrelski et al., 1993; Yoneda et al., 2017). However, Roy and Gupta (2004) reported that the exclusion of denaturing factors can reverse denatured proteins to their native states. Prestrelski et al. (1993) reported that structural modifications such as protein unfolding by dehydration can be fully reversed by rehydration.
We hypothesized that the freeze-dried pork powder could be used for comminuted meat products with the recovery of protein functionality by rehydration, and the processing characteristics of freeze-dried pork powder might be similar to those of freeze-thawed pork. Therefore, the pork emulsions were prepared using freeze-dried pork powder with two levels of sodium chloride. The gelling and emulsifying properties were measured and compared to those of pork emulsions manufactured from freeze-thawed pork.
Materials and Methods
Front legs from three female pig carcasses were purchased from a local meat shop two days after slaughter. Pork from each carcass was allocated to each batch (a total of three batches). The pork was cut into cubes (approximately 3 cm×3 cm×3 cm), and then the pork from each carcass was divided into two groups (freeze-thawed and freeze-dried groups) and vacuum packed. One group of packaged pork was frozen at –20°C, while the other group was lyophilized after two days of freezing. The pork was freeze-dried at a chamber pressure of 5 mTorr at room temperature using a freeze dryer (Bondiro, Ilshin, Seoul, Korea) for 7 days, followed by pulverization and storage at –20°C until use. The frozen pork was left at –20°C for 30 days and then thawed at 4°C for 48 h before use. All the experiments were conducted in three independent batches.
Three meat samples were collected from each treatment batch. The moisture and crude protein contents were measured by method 981.10 of AOAC (2010). The crude protein contents of the freeze-thawed and freeze-dried powder samples were 19.72% and 70.10%, respectively. The freeze-dried pork powder samples were rehydrated with distilled water overnight at 4°C to obtain the same crude protein content as that of freeze-thawed pork before all analyses.
Myofibrillar proteins were extracted according to the method described by Lee et al. (2021b). The sample (3 g) was homogenized twice with cold isolation buffer (2 mM MgCl2, 1 mM ethylene glycol tetraacetic acid [EGTA] and 0.1 M KCl dissolved in 10 mM potassium phosphate [pH 7.0]) at 12,000 rpm for 15 s with a homogenizer (T25 basic, IKA GmbH & Co. KG, Germany). The homogenate was filtered through a mesh strainer (1.0 mm) to remove connective tissues and then centrifuged at 2,000×g for 10 min at 4°C (1580R, LABOGENE, Lynge, Denmark). The pellets were washed with isolation buffer (20 mL) and centrifuged twice under the same conditions as described above. Then, an aliquot (20 mL) of 100 mM NaCl was added to the pellet and washing was performed three times. The final pellets were vortexed with 0.1 M potassium phosphate buffer (pH 7.4) to make a myofibril suspension.
The content of bound bromophenol blue (BPB) of the myofibrillar proteins (protein concentration of 1 mg/mL) was used to determine the surface hydrophobicity of the samples according to the method described by Chen et al. (2015). The samples that were reacted with BPB were centrifuged, and the supernatants were diluted 10-fold. Phosphate buffer and BPB were reacted together to set up the control sample. The absorbance of the diluent was determined at 595 nm using a plate reader (Varioskan LUX, Thermo Fisher Scientific, MA, USA). The bound BPB content was calculated using the following equation:
The extracted myofibrillar proteins were dissolved in 0.6 M NaCl solution and diluted to obtain a protein concentration of 0.5 mg/mL. The samples were then scanned using a fluorescence spectrophotometer (Varioskan LUX) following the methods of Lee et al. (2021a). The fluorescence intensity was obtained between 300 and 400 nm at an excitation wavelength of 280 nm and a scanning speed of 1,000 nm/min. The excitation and emission slit widths were 5 nm.
The carbonyl and total free sulfhydryl groups were monitored to determine the protein oxidation. Carbonyl content was evaluated by measuring the absorbance of the derivatized samples with dinitrophenylhydrazine (DNPH) at 370 nm (Varioskan LUX) in accordance with the protocol of Chen et al. (2015). Protein content was monitored using a Bio-Rad protein assay (#5000006; Bio-Rad Laboratories, Richmond, CA, USA) at an absorbance of 595 nm, and bovine serum albumin was used to generate the standard curve. A molar absorptivity of 21.0 nM−1 cm−1 was used to express the carbonyl content as nmol/mg of protein.
The total free sulfhydryl group content was determined in the myofibrillar protein extract (protein concentration of 1 mg/mL) using the method described by Hofmann and Hamm (1978). The myofibril suspension was reacted with 5,5'-dithiobis [2-nitrobenzoic acid] (DTNB) solution, and the absorbance of the mixture was measured at 412 nm using a plate reader (Varioskan LUX). A molar absorptivity of 11,400 M−1 cm−1 was used to calculate the amount of total free sulfhydryl groups.
The solubility of total and sarcoplasmic proteins was measured according to the method previously described by Chan et al. (2011). For total protein solubility, the pork sample (1 g) was homogenized with 40 mL of 0.55 M potassium iodide in 0.05 M phosphate buffer (pH 7.4) and centrifuged at 2,000×g for 10 min (1580R, LABOGENE). For sarcoplasmic protein solubility, pork samples (2 g) were homogenized (T25 basic, IKA GmbH & Co.) with 40 mL of 0.03 M phosphate buffer (pH 7.4) and then centrifuged (1580R, LABOGENE). The filtrate of each supernatant (No. 1, Whatman, Maidstone, UK) was analyzed for protein concentration using a Bio-Rad assay (#5000006, Bio-Rad Laboratories). The solubility of myofibrillar protein was calculated by subtracting the sarcoplasmic protein solubility from the total protein solubility.
The content of non-heme iron was determined using the Ferrozine method according to the method described by Pourkhalili et al. (2013). The pork sample (0.5 g) was dissolved in 1 mL of 2% ascorbic acid in 0.2 M HCl and 3 mL of 0.1 M citrate phosphate buffer (pH 5.5) and reacted at 25°C for 15 min. After adding 2 mL of 11.3% trichloroacetic acid (TCA), the mixture was centrifuged at 2,090×g for 10 min (1580R, LABOGENE). An aliquot (2 mL) of the supernatant was reacted with 10% ammonium acetate (0.8 mL) and ferrozine reagent (0.2 mL). The absorbance was read at 562 nm using a plate reader (Varioskan LUX), and the standard curve was made with FeCl3 (1 g/L stock solution).
Myosin was extracted using a method previously reported by Fu et al. (2020). Pork samples (1 g) were homogenized (T25 basic, IKA GmbH & Co.) with 10 mL of isolation buffer (2 mM MgCl2, 1 mM EGTA, and 0.1 M KCl dissolved in 10 mM potassium phosphate [pH 7.0]) and centrifuged at 2,000×g for 10 min (1580R, LABOGENE). An aliquot (5 mL) of Tris buffer (5 mM Tris, 1 mM EGTA, and 1 mM 1,4-dithiothreitol [DTT], pH 7.8) was added to the pellet, homogenized, and centrifuged again. The pellet was homogenized with 150 mM ethylenediaminetetraacetic acid (EDTA) in 150 mM potassium phosphate (pH 6.5), and the mixture was centrifuged. G-S buffer (0.4 M NaCl, 5 mM MgCl2, 5 mM Na4P2O7, and 150 mM Na3PO4, pH 6.0) was stirred with the pellet for 20 min, followed by centrifugation. The pellet was collected and the supernatant was filtered through a 1.0 mm mesh strainer. Distilled water was added to both the pellet and the filtrate of the supernatant, followed by stirring for 10 min and centrifugation. Each pellet was treated with 0.6 M KCl dissolved in 40 mM potassium phosphate buffer (pH 6.5) and diluted to the protein concentration of 0.5 mg/mL.
Polyacrylamide gel (12.5%) was used to perform sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (Lee et al., 2020). The extracted myosin samples (protein concentration, 0.5 mg/mL) were diluted twice (v/v) with sample buffer (EBA-1051, Elpis Biotech, Daejeon, Korea) and heated at 95°C. The samples (0.5 mg protein/mL) and protein ladder (3454A, Takara Bio, Shiga, Japan) were loaded at 10 and 5 μL, respectively. The bands were stained with a solution containing Coomassie brilliant blue, acetic acid, and methanol overnight. Then, a buffer containing acetic acid and methanol was used to de-stain the gels. The gels were scanned at an optical resolution (63.5 μm/pixel) with a densitometer (GS-710, Bio-Rad Laboratories) and analyzed using Image Master 2D Platinum v5.0 (GE Healthcare, formerly Amersham Biosciences, Seoul, Korea).
The rehydrated pork powder with distilled water exhibiting the same protein content as that of the freeze-thawed pork was employed for the production of meat batter. The meat batter was manufactured with two sodium chloride levels with both freeze-thawed pork (emulsion gel manufactured with freeze-thawed pork, GFT) and rehydrated pork powder (emulsion gel manufactured with freeze-dried pork powder, GFD). The main materials for the meat batter include pork (70%), back fat (20%), ice (5%), and cold water (5%). The additives were used in the following proportions of the main materials: 1.5% and 2.0% sodium chloride, 0.3% sodium pyrophosphate, 0.01% NaNO2, and 0.03% ascorbic acid. The mixture without back fat was ground in a food processor (FPM250, De’Longhi-Kenwood Appliances, Treviso, Italy) for 2 min, and after the addition of back fat, the homogenate was ground again for 2 min. The meat batter (50 g) was stuffed in a steel can (5 cm in diameter) and vacuum-packed. The steel was then heated at 80°C until it reached the core temperature of 75°C, followed by cooling at 25°C. Five gels prepared in each treatment/batch were randomly allocated for quality analysis, and three gels were stored at –50°C for subsequent analysis.
The cooking losses of the emulsion gels were calculated as the ratio of the emulsion batter before and after cooking using the following equation:
The textural properties were monitored using a two-bite system with a texture analyzer (A-XT2, Stable Micro Systems, UK) using a compression probe (70 mm). The gels were cut into 2 cm×2 cm×1.5 cm and compressed (70%) at a test speed of 2 mm/s. Hardness (N) was used to determine the strength of the gels.
The malondialdehyde (MDA) content was measured to monitor the lipid oxidation of the gels. The sample homogenate in distilled water and 7.2% butylated hydroxytoluene (BHT) was mixed with 6 M NaOH at 60°C for 45 min. After adding 1 mL of acetonitrile, the mixture was vortexed for 10 s and centrifuged. The supernatant was filtered through a syringe filter (Whatman PLC) and analyzed by high-performance liquid chromatography (HPLC) following the method of Jung et al. (2016). The MDA content of the samples was expressed as mg MDA/kg meat.
The color (CIE L*, a*, and b*) of the gels was measured at two different parts of the cut inner surface using a colorimeter (CM-350d, Konica Minolta, Tokyo, Japan) with an illuminant D65, illumination area of 30 mm, and standard observer of 10°, and the average value was considered. The results were automatically analyzed using Spectra Magic Software (SpectramagicTM NX, Konica Minolta).
The total pigment and nitrosyl-hemochrome contents of the gels were measured by extraction with acetone (Lee et al., 2018). For determining total pigment, the sample (10 g) was mixed with 2 mL of distilled water, 1 mL of HCl, and 40 mL of 80% acetone and kept in the dark at 4°C for 1 h. After filtering the supernatant through filter paper (No. 1, Whatman), the absorbance of the filtrate was measured at 640 nm (Varioskan LUX). The concentration of the total pigment (ppm) was calculated by multiplying the absorbance by 680.
The samples for determining nitrosyl-hemochrome were mixed with 3 mL of distilled water and 40 mL of 80% acetone and kept in the dark for 5 min. The supernatant was filtered through filter paper (No. 1, Whatman), and its absorbance was recorded at 540 nm (Varioskan LUX). The concentration of nitrosyl-hemochrome (ppm) was calculated by multiplying the absorbance by 290. The nitrosyl-hemochrome content was expressed using the following formula:
Three iterations were performed (three batches with three samples/treatment/batch), and the results were analyzed using a mixed model with a randomized complete block design (where a batch is a block). The results were expressed as the least-square mean and standard error of the least-square means, and the significance of the main effects was tested using Tukey's multiple tests, with p<0.05 considered statistically significant. Statistical analysis was performed using SAS version 9.3, SAS Institute, Cary, NC, USA).
Results and Discussion
The ice crystals sublimate during primary drying (Tang and Pikal, 2004). The moisture content of the samples after primary drying appears to be as high as 7%–8% (Severo et al., 2013). The remaining water during secondary drying is generally bound to proteins strongly. After the removal of bound water, only 1%–2% of the moisture remained in the samples following secondary drying (Severo et al., 2013). In this study, the moisture content of the freeze-dried powder was 1.34% (Table 1). Thus, freeze-drying of pork appeared to be completed successfully through both primary and secondary drying.
| Treatment | SEM1) | ||
|---|---|---|---|
| Freeze-thawed pork | Freeze-dried pork powder | ||
| Moisture (%) | 73.77A | 1.34B | 0.179 |
| Crude protein (%) | 19.72B | 70.10A | 1.035 |
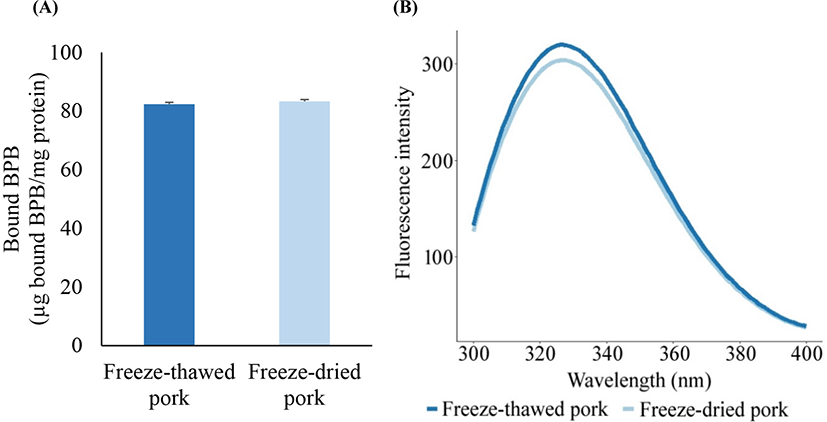
The unfolding of proteins changes their tertiary structure, resulting in an increase in surface hydrophobicity and a decrease in tryptophan fluorescence intensity upon exposure to amino residues that were originally buried in the native structure (Deng et al., 2015). The increase in surface hydrophobicity and the consequent aggregation among unfolded proteins negatively affect protein solubility, which is an important property in the quality of gelling and emulsifying agents (Careche et al., 1998; Dissanayake and Vasiljevic, 2009). The surface hydrophobicity of myofibrillar proteins extracted from the rehydrated pork powder did not differ from that of the freeze-thawed pork (Fig. 1A). In addition, no significant difference in fluorescence intensity was observed between the two groups (Fig. 1B). Proteins that are highly stable during dehydration, can either maintain the folded structure during rehydration or refold to their native structure, despite unfolding during dehydration (Prestrelski et al., 1993; Roy and Gupta, 2004). The removal of water from proteins by freeze-drying reconstitutes proteins through the aggregation of unfolded proteins; thus, modified proteins in the structure are degraded during the drying process (Ohtake et al., 2011). The aggregation or degradation of meat proteins changes the surface hydrophobicity and tryptophan fluorescence intensity contrastingly (Lee et al., 2021a; Masniyom and Benjakul, 2004). However, the refolded proteins during rehydration might not induce any change in the tertiary structure of the freeze-dried pork powder compared to the freeze-thawed pork.
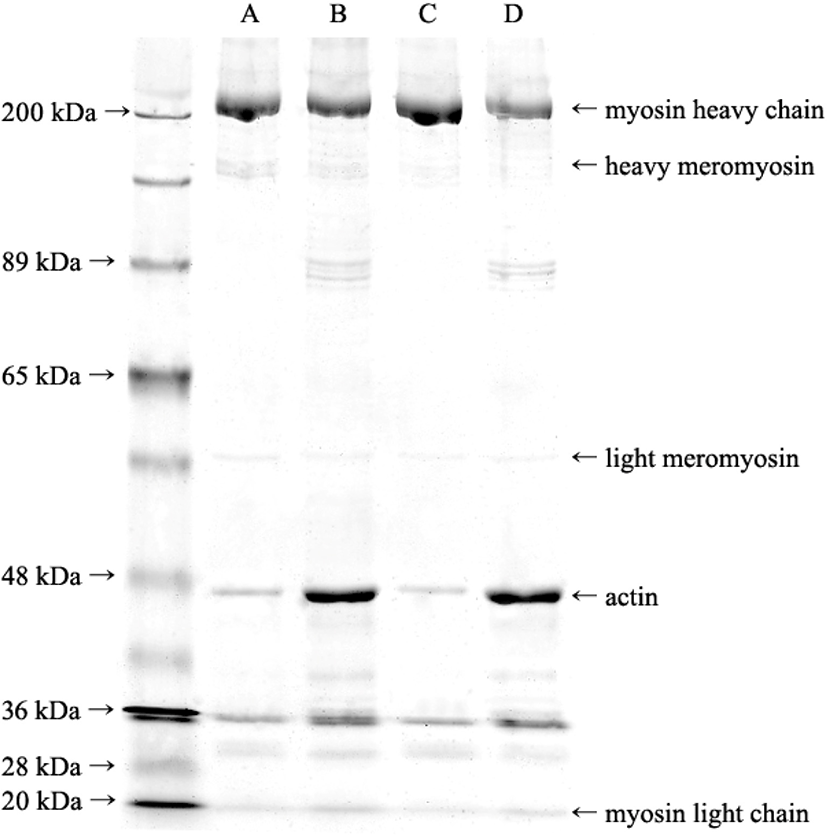
The protein carbonyl content in the rehydrated pork powder was significantly higher than that in the freeze-thawed pork (p<0.05, Table 2). Myofibrillar proteins in frozen meat are prone to high oxidation because more amino acid residues are exposed to the frozen concentrate of solutes, including oxidants (Utrera et al., 2014). The frozen meat temperature in the chamber is increased with ice sublimation during primary drying in a freeze-dryer. This increased temperature in the presence of water is favorable for protein oxidation (Tang and Pikal, 2004). Therefore, in this study, protein oxidation may have occurred during freeze-drying. Generally, protein oxidation results in the aggregation of proteins by disulfide bonds, leading to a decrease in both the tryptophan fluorescence intensity and the total free sulfhydryl group content of protein (Utrera et al., 2014). However, there was no difference in tryptophan fluorescence intensity between the freeze-thawed pork and freeze-dried pork. Furthermore, the free sulfhydryl group content was higher in freeze-dried pork than in freeze-thawed pork (p<0.05, Table 2). This result is in agreement with the increase in free sulfhydryl groups found in myosin from freeze-dried squid when compared to raw squid because of myosin degradation during the drying process (Deng et al., 2015). Therefore, degradation and aggregation may occur in pork proteins during freeze-drying. This result was consistent with the SDS-PAGE results (Fig. 2). In this study, we observed the electrophoretogram of SDS-PAGE in the supernatant (myosin) and pellet (myofibrillar proteins) of myosin extract (Fig. 2). The bands for light meromyosin and myosin light and heavy chains appear to have similar intensities for freeze-thawed and freeze-dried pork. However, the band intensity of heavy meromyosin in freeze-dried pork was lower than that in freeze-dried pork. In fact, the heavy meromyosin bands for freeze-dried pork appeared very faintly on the gel, indicating that meromyosin was denatured and degraded during freeze-drying. Li and Xiong (2015) have shown that the relative intensity of heavy meromyosin is reduced with increasing hydrogen peroxide dosage. Thus, the myosin rod containing subfragment-2 of heavy meromyosin might be attenuated by increasing oxidative stress.
In this study, we discovered that the solubilities of total proteins, sarcoplasmic proteins, and myofibrillar proteins were significantly lower in the rehydrated pork powder than in the freeze-thawed pork (Table 2). This result was related to the protein oxidation in freeze-dried pork. Protein oxidation is a detrimental factor for the functional properties of proteins, such as, decreased solubility due to the formation of carbonyl groups and aggregation (Zhang et al., 2013). Prestrelski et al. (1993) found that the aggregation and precipitation of freeze-dried proteins exhibited irreversible conformational changes caused by freeze-drying when they were rehydrated. Moreover, precipitation of denatured myofibrillar proteins lowers protein solubility by affecting the extractability of proteins by encasing the myosin heavy chain (Chen et al., 2015).
A previous study reported the denaturation of myoglobin in freeze-dried meat (King and Chen, 1998). Myoglobin is a heme protein that contains iron ions, and heme iron is generally converted into non-heme iron by the denaturation of myoglobin (Lombardi-Boccia et al., 2002). Non-heme iron is a type of oxidation catalyst used in meat products. However, in this study, there was no significant difference in non-heme iron content between the two treatments.
The emulsion gel was manufactured using freeze-thawed pork or freeze-dried pork powder with either 1.5% or 2.0% NaCl. GFD showed higher cooking loss than GFT when using 2.0% NaCl (p<0.05, Table 3). This high cooking loss of the emulsion gel is attributed to the low retention of water and fat in the gel, as the retention is affected by the water-holding capacity of proteins, amount of solubilized protein, and structural integrity of the gel (Sun and Holley, 2011). Chen et al. (2016) reported that a strand-type gel with low integrity was formed when the solubilization of myofibrillar protein was low, while an aggregated-type gel with high integrity was obtained with an increase in protein solubilization. Therefore, the lower water and fat retention in GFD compared to GFT was caused by the lower gel integrity due to the lower myofibrillar protein solubility in the freeze-dried pork powder. In addition, the freeze-dried pork powder contained a higher carbonyl group content than the freeze-thawed pork. Protein carbonyls form cross-linkages through condensation with neighboring amino acids; further, the loss of amino groups in side chains disrupts the electrostatic arrangement (Estévez et al., 2011). Electrostatic repulsion among proteins is a major driving factor contributing to the filament lattice spacing, which is a key parameter determining the water retention of muscle fibers. Since the protein gel swells during water absorption because of electrostatic repulsion among proteins, the reduction of these forces by protein aggregation can lower the barrier against moisture loss (Kristinsson and Hultin, 2003). Accordingly, in this study, we identified that the occurrence of protein aggregation during freeze-drying might induce an increase in cooking loss in GFD. Although the freeze-dried pork powder offered low protein solubility, the cooking loss of the gel was not different between GFT and GFD with 1.5% NaCl (Table 3). Hence, a salt level of 1.5% might result in no differences in cooking loss between both treatments; moreover, the protein extractability is low at this concentration. Barbut (1995) reported that 1.5% NaCl is the threshold salt level for the water-holding capacity of meat batter, and increasing the salt concentration from 1.5% to 2.5% doubled the amount of extractable proteins. This is because NaCl swells the myofilament in muscle fibers, thereby enlarging the space to retain moisture in the muscle protein, and improving the water retention as well as increasing the myofibrillar protein solubility (Ruusunen and Puolanne, 2005). Therefore, the cooking loss for both treatments in this study was lower in the 2.0% NaCl group than in the 1.5% NaCl group (p<0.05).
| Treatment Pork | NaCl level | Properties | |
|---|---|---|---|
| Cooking loss (%) | Hardness (N) | ||
| GFT | 1.5% | 7.47A | 78.59B |
| GFD | 7.60A | 48.25C | |
| GFT | 2.0% | 5.23C | 100.84A |
| GFD | 6.23B | 55.55C | |
| SEM1) | 0.186 | 1.889 | |
GFD had a significantly lower hardness (values of 48.25 and 55.55 N) than GFT (values of 78.59 and 100.84 N) at salt levels of 1.5% and 2.5%, respectively (p<0.05, Table 3). The lower protein solubility of the freeze-dried pork powder might contribute to weakening the emulsion gels because solubilized proteins form a gel network through interactions with insoluble proteins, and also by holding water molecules and fat particles (Choi et al., 2020; Sun and Holley, 2011). An increase in salt levels results in an increase in the myofibrillar protein solubility due to enhanced electrical repulsion (Chen et al., 2016). However, the hardness of GFD did not significantly change with increasing NaCl concentration, whereas that of GFT increased at 2.0% NaCl. Myofibrillar proteins are the major proteins responsible for the texture of meat products. Myosin is the most important component in the gelation of myofibrillar proteins. The gelation of muscle proteins leads to the formation of a three-dimensional network system with a helix-coil transition, resulting in the formation of disulfides in protein molecule tails (Sun and Holley, 2011). Samejima et al. (1981) reported that two components of myosin, subfragment-1 and the rod, formed a heat-induced gel. The authors reported that myosin binds head-to-head and self-associates by cross-linking the tail. Heavy and light meromyosin form a gel by irreversible associations and reversible helix-coil transitions, respectively (Ishioroshi et al., 1982). We observed protein oxidation in the freeze-dried pork powder, which contained a higher carbonyl group content, than in freeze-thawed pork (Table 2). Moreover, the electrophoretogram of SDS-PAGE showed that myosin was denatured and degraded after freeze-drying (Fig. 2). Therefore, the oxidative damage to myosin might have decreased the protein solubility and gel hardness, because protein denaturation before heating hinders gelation.
Color analysis yielded lower a* and higher b* values in GFD than in GFT (p<0.05; Table 4). However, the nitrosyl-hemochrome content was not significantly different among the treatments employed (p>0.05).
In this study, the proteins in the freeze-dried pork powder were oxidized; consequently, the solubility of sarcoplasmic protein, consisting of myoglobin, was lower in the powder than in the thawed pork (Table 2). Myoglobin comprises a heme group and a globin, and its autoxidation is typically related to the oxidation of an iron atom in the heme group instead of the oxidation of the globin chain (Wang et al., 2019). The conversion of myoglobin to metmyoglobin during the oxidation of iron atoms can result in different meat colors (Faustman et al., 2010). Although there were no significant changes in the non-heme iron content between the powder and thawed pork, the myoglobin seemed to be irreversibly denatured by freeze-drying. Moorthy et al. (2014) reported that lyophilized myoglobin underwent significant aggregation during storage. Moreover, King and Chen (1998) reported that the freeze-drying of beef and pork decreased the myoglobin content. The authors explained that freeze-dried powder can contain degraded myoglobin by the enhanced oxidation due to the higher porosity caused by the sublimation of ice crystals. Therefore, dehydration during freeze-drying might result in the aggregation of myoglobin.
Lipid oxidation can also lower the redox stability of myoglobin (Faustman et al., 2010). In this study, the MDA content in GFD was significantly higher than that in GFT (p<0.05; Table 5). MDA, which is a secondary product of lipid oxidation, can directly cause the autoxidation of myoglobin. This occurs by destruction of the heme pocket formed by the compact globin via reaction with aldehydes, which are the byproducts of lipid oxidation (Wang et al., 2019). Oxymyoglobin and deoxymyoglobin have higher resistance to heat and are denatured into ferrohemochrome, which is a red globin hemochromogen. On the other hand, metmyoglobin has lower resistance to heat and, when heated, denatures into ferrihemochrome, which is a brown globin hemichromogen (Jo et al., 2020a). Therefore, in this study, the denaturation of myoglobin in freeze-dried pork powder might have induced a lower a* value in GFD than in GFT.
| Treatment Pork | NaCl level | Malondialdehyde content (mg/kg) |
|---|---|---|
| GFT | 1.5% | 0.11B |
| GFD | 0.14A | |
| GFT | 2.0% | 0.11B |
| GFD | 0.14A | |
| SEM1) | 0.003 |
NO-myoglobin, which is formed via the combination of nitric oxide and myoglobin, is unstable and converted into the stable pigment nitrosyl-hemochrome when heated (Jung et al., 2017). Metmyoglobin can be transformed to NO-metmyoglobin by reacting with nitric oxide and reducing it to form NO-myoglobin, which contributes to the cured pink color (Jo et al., 2020b). Therefore, the transition of myoglobin to NO-myoglobin appears to mainly form nitrosyl-hemochrome in GFT. However, the reduction of NO-metmyoglobin might have dominantly formed nitrosyl-hemochrome in GFD, as we included ascorbic acid in the meat batter. Therefore, although there was no significant difference in the content of nitrosyl-hemochrome between GFD and GFT, its formation mechanism may differ. This can be inferred from the fact that the two gels exhibited significantly different a* values, and the myoglobin of the freeze-dried pork powder was potentially oxidized and denatured more than in the freeze-thawed pork.
Meanwhile, the higher b* value in GFD might arise due to browning after freeze-drying. Brown pigmentation caused by the non-enzymatic Maillard reaction has been reported to occur in lyophilized muscle food. Although meat contains only a low carbohydrate content, the carbonyl compounds and secondary lipid oxidation products (including aldehydes) that form during storage can participate in the Maillard reaction with amino acids, resulting in browning (Harnkarnsujarit et al., 2015). Moreover, the manifestation of the brown color and dark pigmentation might have also been enhanced in GFD by the autoxidation of myoglobin into metmyoglobin (George and Stratmann, 1952; Jo et al., 2020a).
Conclusion
Freeze-dried pork powder exhibited higher protein oxidation and lower protein solubility than freeze-thawed pork. GFD presented a higher cooking loss and lower hardness than GFT. In addition, decreased redness and increased yellowness were observed in GFD compared to GFT. Hence, we deduced that the denaturation of proteins during freeze-drying may negatively affect the pork powder used as a raw material for comminuted meat products. However, the control of freezing conditions, such as temperature and duration, to determine the protein functionality of the dried meat powder may partially reduce the occurrence of oxidation and denaturation. In addition, the use of stabilizers such as sugars may inhibit the denaturation of proteins during freeze-drying. Therefore, methods to maintain the protein functionality in freeze-dried pork powder should be investigated in future studies to use this powder as a raw material in comminuted meat products.