










Introduction
Colons are culture devices for numerous intestinal microbes. The intestinal microbiota not only protects against the invasion of pathogens but also participates in the immune system as well as in the production of vitamins and short-chain fatty acids (SCFAs) to supply nutrients to regulate human metabolism. Additionally, intestinal microbiota is known to have an influence on human health and various diseases. In fact, the intestinal microbiota of patients with diseases such as constipation is different from that in normal people. Previous studies have shown that the abundance of specific species among intestinal bacteria differs in patients with constipation. People with symptoms of constipation reportedly have decreased abundance of Bifidobacteria and Lactobacillus and increased abundance of Bacteroidetes compared to the general population (D’Onofrio et al., 2020; Wang et al., 2020). Such changes in the intestinal microbiota affect intestinal motility and intestinal environment (Zhao and Yu, 2016). Administration of Bifidobacterium adolescentis to an animal model with loperamide-induced constipation improved constipation and altered the intestinal microbiome composition (Wang et al., 2017).
Several studies have demonstrated that certain probiotics strains could play a beneficial role in relieving constipation symptoms (Bekkali et al., 2007; Koebnick et al., 2003). These probiotics are used to make yogurt. Traditional yogurt starters Streptococcus thermophilus, Lactobacillus bulgaricus, and L. acidophilus are added to Bifidobacterium strains (B. bifidum and B. lactis) to make yogurt. These multiple strains are widely used as starters for yogurt production (Ahn, 2014; Lim et al., 2015). Intake of fermented milk containing multi-strains is effective in improving irritable bowel syndrome, including constipation (Wen et al., 2020).
The effect of probiotics varies depending on the strain (Aloisio et al., 2012; Presti et al., 2015), and using mixed strains may be more efficient than using single strains because of the diversity and complexity of irritable bowel diseases. Moreover, the use of mixed strains improves intestinal adhesion and the production of various metabolites and is more effective in improving intestinal diseases compared to a single strain (Yoon et al., 2014). Furthermore, supplements containing multi-strain probiotics are effective in treating subjects with irritable bowel diseases and improving the composition of the intestinal microbiota (Mezzasalma et al., 2019). However, it is currently unclear whether this is due to synergistic interactions between strains or the higher probiotics doses used in some studies (Chapman et al., 2011). Multi-strain probiotics appear to exhibit greater efficacy than single strains according to a limited number of studies.
In this study, Sprague-Dawley rats with loperamide-induced constipation were orally administered probiotics and prebiotics samples. To evaluate their constipation-relieving effect in rats, changes in the stool parameters, gastrointestinal transit ratio, and intestinal microbiota were analyzed. Combination treatment of synbiotics and probiotics effectively alleviated loperamide-induced constipation.
Materials and Methods
Male Sprague-Dawley rats (6 weeks old, 160–180 g) were purchased from Oriental Bio and allowed to adapt to the environment for 1 week. All experiments were approved by the Korea University Institutional Animal Care & Use Committee (Approval number: KUIACUC-2020-0026). In the breeding environment, the temperature was 21±1°C, the relative humidity was 50%–55%, a 12 h light/dark cycle was maintained, and standard commercial feed and water were supplied ad libitum. A probiotics sample containing a mixture of Lactobacillus plantarum, L. acidophilus, Bifidobacterium bifidum, B. lactis, and Streptococcus thermophilus was obtained from Chong Kun Dang HealthCare (Seoul, Korea). Petri dishes containing strain-specific selective agar were used to count and confirm the number of probiotics. Each rat in acrylic cages was administered either probiotics (0.2 mL of 5.0×109 CFU/g probiotics and prebiotics) or the placebo solution. Loperamide was obtained from Sigma-Aldrich (St. Louis, MO, USA). The prebiotics sample containing a mixture of lactitol (DuPont, Wilmington, MA, USA), Kamut steamed powder (Duri Duri, Nonsan, Korea), and Microbiome X (BioActor, Maastricht, Netherlands) was obtained from Chong Kun Dang Healthcare Other reagents used were general special reagents.
Experimental animals were randomly divided into four groups of six animals each and classified into a control group (Cont), loperamide-induced group (Lop), loperamide-induced group with multi-strain probiotics mixture group (Lop-Pro), and loperamide-induced group with multi-strain probiotics and prebiotics mixture group (Lop-Pro/Pre). In all groups, except for the Cont group, loperamide (3 mg/kg) diluted in physiological saline was administered orally once a day for 6 days to induce constipation. Constipation symptoms were confirmed by measuring the amount of stool.
Cont and Lop groups were orally administered with saline, and the treatment groups (Lop-Pro and Lop-Pro/Pre) were orally administered a multi-strain probiotics (LACTO 5X, 31 mg/kg) or a mixture of multi-strain probiotics and prebiotics (LACTO 5X synbiotics, 31 mg and 120 mg/kg, respectively). All samples were suspended in saline and administered orally once daily for 21 days. The body weight and food intake of all rats were measured twice per week throughout the experiment. All rats were sacrificed by CO2 exposure after 21 days of treatment.
The number, weight, and moisture content of the feces were measured twice in the last week. To examine the fecal moisture content, feces were dried at 70°C for 24 h to measure the dry weight, and the difference in fecal weight before and after drying was divided by the fecal weight and calculated as a percentage.
On day 21, all experimental animals were fasted for 12 h and 1 mL of 8% charcoal was orally administered; 20 min later, the animals were sacrificed to measure the length of the intestine and the distance traveled by the charcoal. Intestinal transit time was calculated using the following equation:
On the day of sacrifice, the colon tissue was excised, cut into cells, fixed in 10% neutral formalin for 18 h or more, dehydrated, paraffin embedded, and prepared into 3–4 μm paraffin sections. Then, hematoxylin and eosin (Sigma-Aldrich) staining was performed and changes in the intestinal membrane thickness were observed under a light microscope (Axio Zoom v.16; Carl Zeiss, Göttingen, Germany). For mucin staining, Alcian blue was used. Additionally, the morphology of the Alcian blue-stained crypt cells in the large intestine was observed using an optical microscope and the Leica Application Suite software (Leica Microsystems, Heerbrugg, Switzerland). Staining of intestinal mucosa cells was observed using an optical microscope and Leica Application Suite software (Leica Microsystems). Analysis of stained mucins was performed using MATLAB software by selecting 10 random cryptic cells from at least 5 fields of view per sample.
Immediately after sacrificing the experimental animals, both sides of the large intestine from the post-cecum to the rectum were removed. The extracted colonic tube was fixed with 10% formaldehyde, subjected to a tissue treatment process, and embedded in paraffin to prepare 5 μm thick sections. The sectioned tissues were deparaffinized with xylene, rehydrated for 5 min each in decreasing ethanol concentrations (100%, 90%, 80%, and 70%), and then stained using c-kit (Santa Cruz; SC-168, Dallas, TX, USA) and primary antibodies. Then, the samples were washed with running water, dehydrated for 5 min each in increasing ethanol concentrations (70%, 80%, 90%, and 100%), washed with xylene, and then sealed. The stained intestinal membrane cells were observed using an optical microscope (MM-400, Nikon, Tokyo, Japan) and analyzed using MATLAB. The number of pixels with RGB values in the stained intestinal membrane cells was observed using an optical microscope (MM-400, Nikon, Tokyo, Japan) and analyzed using MATLAB as follows.
For SCFA analysis, 0.5 g of the cecum content was vortexed after adding 0.5 mL of 90% methanol, centrifuged at 8,000×g for 20 min at 4°C, and the supernatant was filtered through a 0.45 μm Millipore filter (Millipore, Burlington, VT, USA). The SCFA in the filtrate was analyzed using a gas chromatography (YL-6100 GC system, Yong-Lin, Anyang, Korea) equipped with a DB-FRAP 123-3253 column (50 m×0.32 mm×0.5 μM), a flame ionization detector, and an autosampler (HT 300, Young-Lin). The injection volume of the sample was 1 μL, the temperature at the injection port and detector was 200°C and 240°C, respectively, and the analysis conditions were similar to those described by Demigné and Rémésy (1985).
To extract microbial genomic DNA from the intestine of Sprague-Dawley rats subjected to different treatments, cecal contents were collected. The genomic DNA of microorganisms contained in the cecal contents was extracted using the ZR Fecal DNA Kit™ (Zymo Research, Orange County, CA, USA), and the changes in intestinal microorganisms were analyzed using 16S rRNA gene pyrosequencing method (Kim et al., 2012). The nucleotide sequence obtained through pyrosequencing were assigned to operational taxonomic unit (OTU) to obtain the OTU values, and the species with 97% sequence similarities were identified using the CLcommunityTM CD-HIT program (ChunLab, Seoul, Korea) (Li and Godzik, 2006). Taxonomic ranking and classification were classified according to the cut-off criteria and the significant difference between groups was performed using the Kruskal-Wallis test method (p<0.05). Database and the sequencing reads of the 16S rRNA gene from this study were deposited in the EzBioCloud database (ChunLab).
Results and Discussion
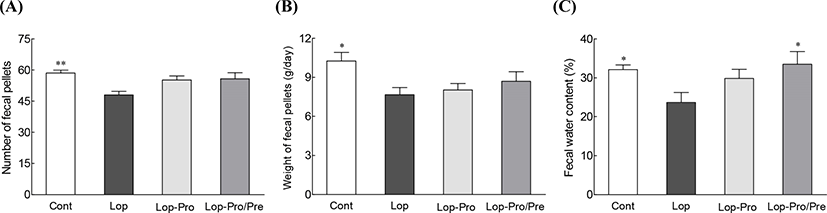
Weight gain, dietary intake, and organ weight were not significantly different between the groups during the experimental period (data not shown). The number and weight of fecal pellets and fecal water content were measured twice prior to sacrifice (Fig. 1) and were significantly different between the Cont and the Lop groups (p<0.01, p<0.05, and p<0.05, respectively). This finding confirmed that loperamide administration induced constipation. Interestingly, the fecal water content was significantly higher in the Lop-Pro/Pre group (33.5%) compared to that of the Lop group (23.7%, p<0.05). From the result of the fecal parameter analysis, we noted that synbiotics administration showed greater improvement in constipation symptoms than administration of multi-strain probiotics alone. Multi-strain probiotics may be more efficient than single-strain probiotics by inducing changes in the diversity of intestinal microbiota, particularly by improving intestinal adhesion and producing various metabolites (Yoon et al., 2014). In this study, a mixture of two Lactobacillus species, one Streptococcus species, two Bifidobacterium species, and a prebiotics material were co-administered to assess constipation-relieving effect. Loperamide is used to induce constipation, inhibit bowel movement, and increase intestinal water absorption (Read, 1983; Théodorou et al., 1991). During constipation, the excretion of fecal pellets significantly decreases along with the water content in the fecal pellets (Wintola et al., 2010; Wu et al., 2011). However, we found that multi-strain probiotics administration caused changes in the fecal number, fecal weight, and fecal moisture content. In particular, the Lop-Pro/Pre group showed a significant increase in fecal pellet moisture content compared to that in the Lop group (Fig. 1).
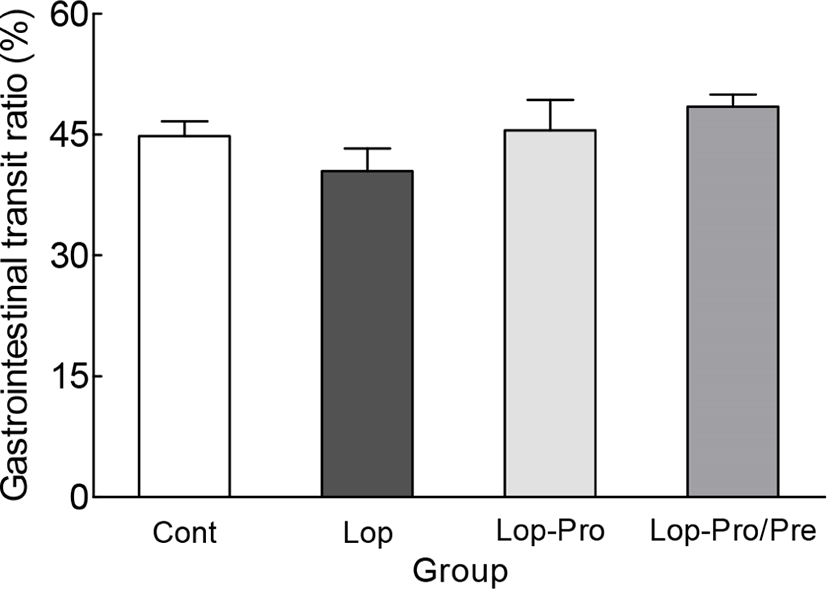
The intestinal transit ratio was measured using activated carbon prior to sacrifice (Fig. 2). The intestinal transit ratio of the Lop group was 40.5%, while that of the Cont group was 44.8%, showing no significant difference. In addition, after induction of constipation, intestinal migration rates of the Lop-Pro and Lop-Pro/Pre groups increased to 45.6% and 48.5%, respectively, but were not significantly different compared to that of the Lop group. Furthermore, the intestinal transit ratio of the Lop-Pro and Lop-Pro/Pre groups increased, but the difference was not significant when compared with that of the Lop group. The reason there was no significant difference in the gastrointestinal transit ratio analysis was that the intestinal length in the Lop group tended to be shorter than in other groups, and there was no statistical difference between the experimental groups.
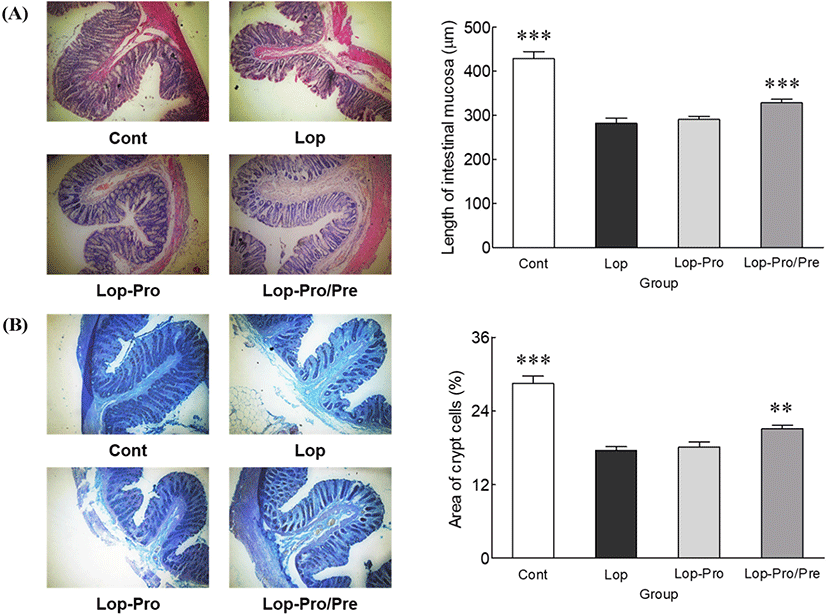
The thickness of the intestinal mucosa was observed using hematoxylin and eosin staining. The mucosal thickness of the Lop group was significantly lower than that of the Cont group (Fig. 3A, p<0.001). After induction of constipation, the intestinal mucosal thickness of the Lop-Pro/Pre group was significantly higher compared to that of the Lop group (16.4%, p<0.001). On the other hand, the Lop-Pro group showed no difference in the mucosal thickness compared to that of the Lop group. The area of mucin-secreting cells, crypt cells, was observed using an optical microscope after Alcian blue staining (Fig. 3B). The area of crypt cells was significantly smaller in the Lop group than in the Cont group (p<0.001). Conversely, crypt cell area considerably increased in the Lop-Pro group compared to the Lop group. In the Lop-Pro/Pre group (21.1%), the crypt cell area increased significantly by 20.6% compared to that of the Lop group (17.5%, p<0.01) and by 16.6% compared to that of the Lop-Pro group (18.1%, p<0.01). Changes in the mucous membrane thickness and the crypt cell area exerted a synergistic effect on improving the intestinal tissue when probiotics and prebiotics were co-administered rather than administration of probiotics alone.
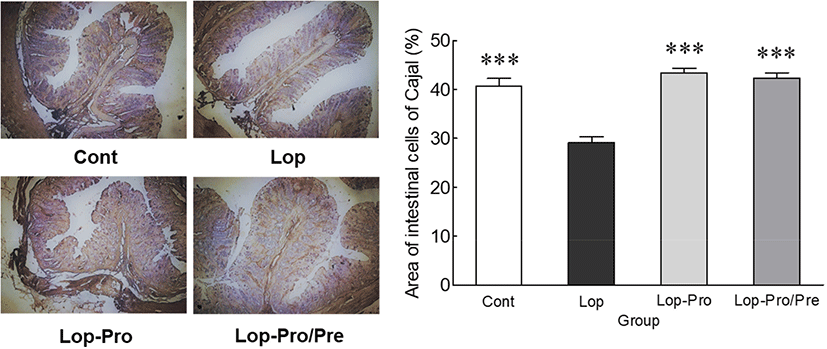
The ICC area, which is related to intestinal peristalsis, was observed using IHC staining (Fig. 4). The ICC area was significantly different between the Lop and Cont groups. Additionally, the ICC area of the Lop-Pro (42.4%) and Lop-Pro/Pre (42.3%) groups significantly increased by 45.4% and 44.8%, respectively, compared to that of the Lop group (29.2%, p<0.001 and p<0.001, respectively). Administration of probiotics alone or the mixture of probiotics and prebiotics reversed constipation-induced decrease in the ICC area; however, the difference in the ICC area was not significant between the two groups. The findings suggest that the inhibitory effect on constipation-induced reduction of the ICC area may be attributed to probiotics.
Induction of constipation by loperamide inhibits intestinal water secretion, decreases colon mucus, and inhibits colon peristalsis, which in turn delays intestinal transit time and increases fecal excretion time (Neri et al., 2012; Shimotoyodome et al., 2000). Moreover, the colonic mucosa is directly associated with constipation (Yang et al., 2008) in that constipation significantly reduces the number of mucus-producing cells, which act as colon barriers by producing mucins, and colonic mucosal thickness, which is related to colon peristalsis (McCullough et al., 1998). In the Lop group, the intestinal transit ratio (Fig. 2), the colonic mucosal thickness, and mucus-producing cell area (Fig. 3) decreased. Multi-strain probiotics and prebiotics administration (Lop-Pro and Lop-Pro/Pre groups) non-significantly increased the intestinal transit ratio, whereas the intestinal mucosal thickness, crypt cell area, and ICC area were significantly different compared to those of the Lop group. When constipation is induced, the decrease in the number of crypts reduces mucus secretion and delays passage of fluids through the intestine (Jeon et al., 2007; Shimotoyodome et al., 2001). In the present study, combined treatment with multi-strain probiotics and prebiotics significantly increased the regeneration of crypt cells compared to that of the Lop and Lop-Pro groups (Fig. 3B). Probiotics and prebiotics together exert a synergistic effect that can inhibit epithelial crypt cell damage. ICC is a cell that regulates colon peristalsis and is closely associated with constipation (Burns et al., 1997; He et al., 2000). Decreased ICC is related to smooth muscle contraction activity and bowel movements, resulting in constipation where there is difficulty in normal bowel movement (He et al., 2000; Wedel et al., 2002). Our results indicated that administration of multi-strain probiotics showed an ameliorating effect on reduced ICC area that was due to long-term constipation (Fig. 4).
The levels of acetic acid, propionic acid, and butyric acid, which are SCFA that help improve gut health, and the total SCFA content were analyzed using gas chromatography (Fig. 5). The SCFA content in all groups was significantly different compared to the Lop group. The level of acetic acid, which was the most prevalent SCFA in the samples, was highest in the Cont group. On the other hand, acetic acid levels in the Lop-Pro and Lop-Pro/Pre groups were similar. Cont, Lop-Pro, and Lop-Pro/Pre groups exhibited significantly higher acetic acid levels compared to the Lop group (p<0.01, p<0.001, and p<0.01, respectively). The difference in propionic acid and butyric acid levels between these groups was similar to the difference in acetic acid levels. The total SCFA content was significantly elevated by 56.4% and 54.2% in the Lop-Pro (36.9 mM) and Lop-Pro/Pre (36.4 mM) groups, respectively, compared to the Lop group (23.6 mM, p<0.001 and p<0.01, respectively). However, the total SCFA content was not significantly different between the Lop-Pro and Lop-Pro/Pre groups. Administration of probiotics and synbiotics resulted in a significant increase in the total SCFA content in the constipation model.
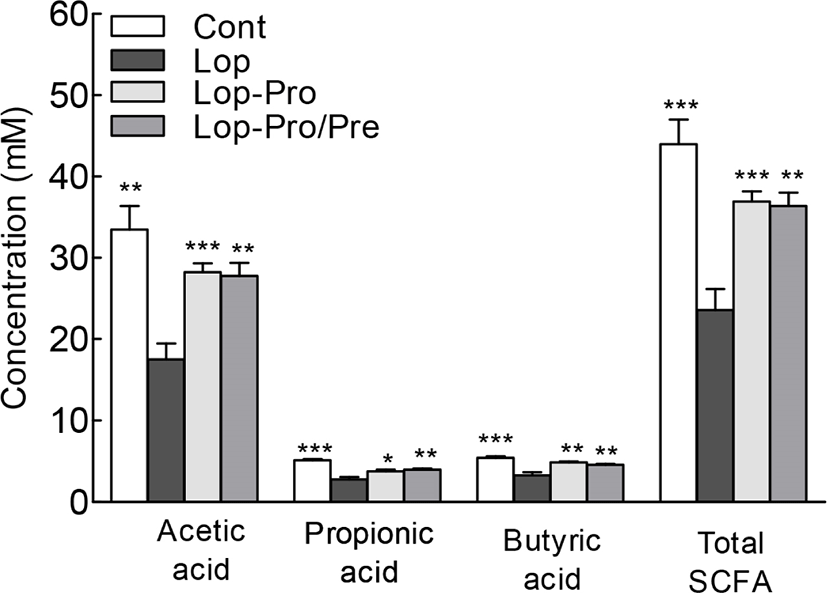
The combination of dietary fiber and probiotics was associated with SCFA production, which is involved in the inhibition of crypt cell loss. As a postbiotic, SCFAs stimulate the proliferation of colon epithelial cells, inhibit the growth of harmful bacteria through acidification of the intestinal environment, and are involved in the integrity of the colon epithelium as a major energy source for intestinal cells (Pruzzo et al., 2000; Topping and Clifton, 2001). Here, the SCFA content was significantly higher in the Lop-Pro and Lop-Pro/Pre groups than in the Lop group (Fig. 5). Postbiotics such as cell-free supernatant, glutathione peroxidase, cell wall fragments, vitamins, phenol-derived metabolites and aromatic amino acids produced by microorganisms are known to have immunomodulatory, anti-inflammatory, antioxidant and anticancer properties (Zolkiewicz et al., 2020). The increase in SCFA content due to the intake of probiotics or synbiotics plays an important role in maintaining bowel health and improving constipation. In addition, the administration of an exopolysaccharide (kefiran; postbiotics) is known to regulate levels of fecal moisture and wet weights of feces (Maeda et al., 2004), and sterilized L. gasseri CP2305 has shown a beneficial improvement in constipation (Sawada et al., 2016).
Changes in intestinal microbiota following multi-strain probiotics administration were analyzed in loperamide-induced constipated rats. Analysis of changes in intestinal microbiota at the phylum level (Fig. 6A) revealed that Firmicutes was the main phylum, occupying a relative abundance ratio of 65.1%–78.4%, and there was no significant difference between the groups. The Lop group showed a decrease in Bacteroidetes abundance and an increase in Verrucomicrobia abundance compared to those of the Cont group. Contrarily, in the Lop-Pro and Lop-Pro/Pre groups, Bacteroidetes abundance increased and that of Verrucomicrobia decreased compared to those of the Lop group. At the order level (Fig. 6B), the abundance of Clostridiales, Bacteroidales, and Lactobacillales decreased in the Lop group but increased in the Lop-Pro and Lop-Pro/Pre groups compared to the Lop group. At the genus level, the relative abundance of Akkermansia was significantly higher in the Lop group than the Cont group (p<0.05, Fig. 6C) and significantly lower in the Lop-Pro group compared to the Lop group (p<0.05). Lactobacillus abundance tended to decrease when constipation was induced, but there was no significant difference between the groups. Furthermore, the relative abundance of Clostridium increased in the Lop group but decreased in the Lop-Pro/Pre group. In particular, the Lop-Pro/Pre group showed a significant difference in the relative abundance of Oscillibacter and Clostridium compared to that of the Lop group (p<0.05). The relative abundance of Bacteroidetes increased in the Lop-Pro and Lop-Pro/Pre groups compared to the Lop group. Although there are differences in the composition of intestinal microbiota, multi-strain probiotics have led to an improvement in the intestinal microbiota.
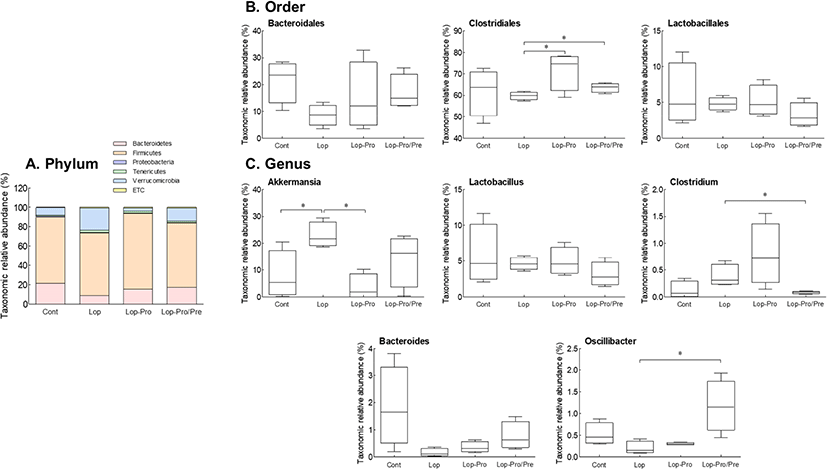
Patients with chronic constipation have relatively lower abundance of beneficial bacteria, such as Lactobacillus, Bifidobacterium, and Bacteroides spp., and greater abundance of potential pathogenic microorganisms, such as Pseudomonas aeruginosa and Campylobacter jejuni, in intestinal microbiota (Gerritsen et al., 2011; Kirgizov et al., 2001). These alterations in intestinal microbiota can affect intestinal motility and the production of metabolites, such as SCFAs, by changing intestinal environment. The main strains of multi-strain probiotics, Bifidobacteria and Lactobacillus, alleviate constipation by producing SCFAs, stimulating intestinal peristalsis, and increasing the water content in fecal pellets (Ojetti et al., 2014). Also, the selected strains used in the experiment have been demonstrated through various animal model experiments and randomized controlled trials to affect the consistency of bowel movements through improvement of the intestinal environment by an increase in beneficial bacteria and metabolites when administered in effective doses (Kaminski et al., 2020; Ohkusa et al., 2019). The use of a mixture of multi-strain probiotics and prebiotics, which is named synbiotics, can be used as a synergistic approach to the survival of probiotics and restore intestinal microbial balance (Khodadad and Sabbaghian, 2010). In addition, reliable evidence has been reported that post-biotics produced through improved intestinal environment and metabolic activity of microorganisms directly or indirectly have beneficial effects on the host (Zolkiewicz et al., 2020). Intestinal microbiota at the phylum level involved Firmicutes, Bacteroidetes, Proteobacteria, and Verrucomicrobia, and these phyla accounted for more than 98% of intestinal microbiota (Guo et al., 2020). During loperamide-induced constipation, the relative abundance of Firmicutes decreases and that of Verrucomicrobia increases (Wang et al., 2020). L. rhamnosus CCFM1068 administration has shown to decrease the abundance of the phylum Verrucomicrobia. The ability of L. rhamnosus CCFM1068 to alleviate constipation symptoms was associated with a decreased abundance of Verrucomicrobia (Wang et al., 2020). We found that multi-strain probiotics and synbiotic co-administration significantly decreased the abundance of Verrucomicrobia compared to that of the Lop group, and constipation-relieving effect may be due to the reduction of the Verrucomicrobia phylum (Fig. 6).
When constipation is induced by loperamide, a decrease in Clostridiales and Lactobacillales abundance and an increase in Bacteroidales abundance is noted (Deng et al., 2018); however, in patients with constipation, Bacteroidales abundance decreases, demonstrating contradictory results (Guo et al., 2020). As shown in Fig. 6B, the decrease in Bacteroidales was confirmed following loperamide treatment, and the levels of the orders Bacteroidales, Clostridiales, and Lactobacillales, which showed changes during constipation induction, were improved when multi-strain probiotics were administered alone or mixed with prebiotics. In particular, in the order Clostridiales, significant differences were observed in the multi-strain probiotics administration groups. In addition, species such as Blatuia, Lachnospira, and Oscillibacter are associated with SCFA production (Zang et al., 2018). In patients with slow-transit constipation, it is inferred that a decrease in SCFA content is associated with a decrease in SCFA-producing microorganisms (Li et al., 2020). The relative abundance of Oscillibacter was significantly increased in the Lop-Pro/Pre group. An increase in the abundance of Akkermansia has been observed in the feces of constipated mice (Wang et al., 2020). An increase in Akkermansia abundance has also been observed in colon cancer patients, and Akkermansia may be related to the disease (Hibberd et al., 2017). As can be seen in Fig. 6C, the relative abundance of Akkermansia was also increased in the Lop group, but the relationship between Akkermansia and constipation should be confirmed. Furthermore, an increase in Clostridium abundance has been reported in children and adults with constipation (Jeffery et al., 2012; Ohara, 2019). As shown in Fig. 6C, the relative abundance of Clostridium in constipated rats was elevated but tended to decrease when multi-strain probiotics and prebiotics were administered. In particular, the relative abundance of Clostridium was significantly reduced in the low-dose group of prebiotics.
The use of probiotics indicated that constipation and intestinal microbiota can be improved. The intestinal pH is lowered by metabolites, such as lactic acid and SCFAs, and this change in the intestinal environment improves intestinal peristalsis and reduces intestinal transit time. It is also involved in bile acid metabolism, changing the shape and concentration of fecal pellets, and activating intestinal movement (Im et al., 2011). The constipation-relieving effect and probiotics activity of yogurt-containing probiotics were improved through modifications such as addition of dietary fiber or various probiotics strains (Jeon and Choi, 2010; Kokke et al., 2008).
Conclusion
We provided experimental evidence that prebiotics/probiotics mixture is an effective approach to changes in cecal microbiome and intestinal health, which relieves constipation. These results were followed by the involvement of prebiotics and probiotics in the processes of alleviating constipation, including improvement of intestinal movement and growth of beneficial intestinal bacteria. According to the results, these effects were mediated by changes in the intestinal mucosal thickness, crypt cell area, and interstitial cells of Cajal area. In addition, changes in Akkermansia, Lactobacillus, Clostridium, Bacteroides and Oscillibacter abundances were involved in the enhancement of the intestinal environment and SCFA production. Collectively, the use of multi-strain probiotics alone (Lop-Pro) showed a constipation-alleviating effect, but synbiotic (Lop-Pro/Pre) used with prebiotics showed better effects in relieving constipation than using probiotics alone.